SCIENCE, VOL. 207, 15 FEBRUARY 1980. pp. 768-769.
Abstract. An abnormal extra band was found on the short arm of the X chromosome in a 46,XY female and in her 46,XY female fetal sibling. Despite presence of the intact Y chromosome, there was no evidence of testicular differentiation in either subject. Production of H-Y antigen was suppressed in both subjects. The data suggest that development of the mammalian testis requires a normal function of the X chromosome.
In mammals, the initially undifferentiated gonad develops as a testis in the presence of the Y chromosome and as an ovary in the absence of the Y chromosome. Yet presence of the Y does not guarantee testicular differentiation, because XY females are known in several mammalian species. Perhaps the most instructive example is the Scandinavian wood lemming, Myopus schisticolor, in which there is a skewed sex ratio with a preponderance of females (1). Roughly half of these female wood lemmings have a male (XY) karyotype, yet they are grossly normal with fertile ovaries. Since XY female wood lemmings are uniformly H-Y negative (2), it might seem at first glance that XY sex reversal in this species is due merely to an abnormality of the Y chromosome. The curious fact is that XY female wood lemmings produce only X-bearing eggs. They do not transmit the Y. Indeed the condition is inherited as an X-linked trait. It appears that a gene on the X chromosome of this species can suppress normal synthesis of H-Y antigen thereby promoting ovarian development in the XY gonadal primordium (3).
Extreme evolutionary conservatism of the mammalian X chromosome (4) implies existence of a similar gene in humans. However, two X chromosomes are needed to support differentiation of the normal human ovary. Whereas X0 gonads of the rodent develop as fertile ovaries, X0 gonads of the human initially develop as ovaries but degenerate, and are represented at around the time of birth by undifferentiated "streaks" devoid of follicles (5). With respect to the number of X chromosomes that are present, XY embryos resemble those with the X0 karyotype. Thus in the functional absence of the testis-determining portion of the Y, the gonads of human XY embryos could be expected to initiate a program of ovarian differentiation, but could not be expected to retain a normal ovarian architecture.
These observations connote a human analog of the wood lemming condition: X-linked suppression of testis-determining H-Y genes in 46,XY phenotypic females with streak gonads (gonadal dysgenesis) (6). In fact there are numerous reports of familial XY gonadal dysgenesis in man. In several of these cases, the pedigree suggests an X-linked mode of inheritance (7). Here we describe failure of testicular differentiation in two 46,XY female sibs, both exhibiting a striking abnormality of the short arm of the X chromosome. The case provides rather more direct evidence of a critical role of the X chromosome in primary sex determination of the mammal in general and of man in particular.
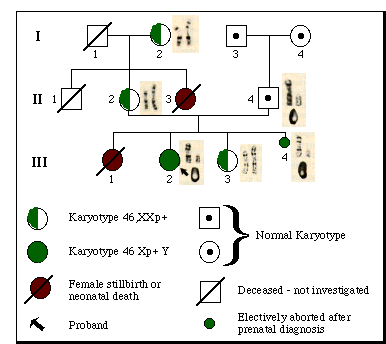
Chromosome studies on a profoundly retarded girl with multiple congenital abnormalities revealed a karyotype of 46,Xp+Y in peripheral blood and skin fibroblast metaphases (Xp+ represents additional material on the short arm of the X; see Fig. 1, III-2). The Giemsa banding pattern of the abnormal X suggested duplication of the p21 and p22 bands, that is, 46,dup(X)(p21--> pter)Y. The Y chromosome was morphologically normal and indistinguishable from the Y chromosome of the father (Fig. 1, II-4) and paternal grandfather (Fig. 1, I-3). Evidently, the aberrant X chromosome was derived from the mother. The proband's maternal grandmother (Fig. 1 I-2), mother (Fig. 1, II-2), and a younger phenotypically normal sister (Fig. 1, III-3) all had a karyotype of 46,XXp+. The proband died at 5 years of age. Autopsy disclosed female internal genitalia and microscopic ovarian remnants consisting of ovarian stroma and degenerating primordial follicles. No testicular tissue was found. Complete details of the case will appear elsewhere (8).
Amniocentesis, performed on the mother during a subsequent pregnancy, revealed cells with a karyotype identical to that of the proband (Fig. 1, III-4). Diagnosis of a grossly abnormal female fetus was confirmed after termination of the pregnancy at 20 weeks of gestation. Multiple anomalies were found similar to those of the proband. Internal genitalia were those of a normal female fetus; normal fetal ovaries were present bilaterally; histological sections failed to reveal any trace of testicular tissue.
Fig. 1. Pedigree showing segregation of the Xp+ chromosome. The Giemsa-banded sex chromosomes and quinacrine-stained Y chromosome are depicted next to the relevant family members. Note the extra band (black) on the abnormal chromosome in comparison with the band on the normal X of the maternal grandmother (I-2), mother (II-2), and normal female sibling (III-3). The extra band is found on the single X chromosome of the XY female proband (III-2) and XY female fetus (III-4).
The following points are worth emphasizing. (i) Serological tests indicated absence of H-Y antigen in skin fibroblasts of the fetus and in blood leukocytes and skin fibroblasts of the proband; (ii) cytogenetic analysis failed to provide evidence for autosome-to-X translocation in any members of the family; and (iii) computer-based videodensitometric analysis of the abnormal chromosome (performed in the Department of Medical Genetics, Mayo Clinic) indicated the likelihood of a duplication of densitometric X bands p12 and 3.
The foregoing observations are consistent with the notion that testicular development is secondary to function of genes on the Y chromosome and on Xp. If X-linked regulatory genes normally served to prevent excess production of H-Y antigen, duplication of these genes might be expected to reduce production of H-Y below a certain critical threshold required for testicular differentiation. Alternatively, the extra bands might interfere with a structural element thereby thwarting production altogether (9). In either event, the result would be subnormal expression of the testis-inducer. The XY gonad would now differentiate as an ovary; the human ovary would degenerate in the absence of the second X. We assume that the disparate gonadal histologies of proband and fetus represent different phases of the same developmental anomaly.
RENÉE BERNSTEIN
Department of Human Genetics,
School of Pathology, South African
Institute for Medical Research, and
University of the Witwatersrand,
Johannesburg, South AfricaGLORIA C. KOO
Memorial Sloan-Kettering Cancer
Center, New York 10021STEPHEN S. WACHTEL
Memorial Sloan-Kettering Cancer
Center and Department of Pediatrics,
New York Hospital -Cornell
Medical Center, New York 10021
14 June 1979
References and Notes
1. K. Fredga, A. Gropp, H. Winking, F. Frank, Nature (London) 261, 225 (1976).
2. S. S. Wachtel, G. C. Koo, S. Ohno, A. Gropp, V. G. Dev, R. Tantravahi, D. A. Miller, O. J. Miller, ibid. 264, 638 (1976).
3. K. Fredga, A. Gropp, H. Winking, F. Frank, Hereditas 85, 101 (1977). More recently these authors identified two different G-banded X chromosomes in the wood lemming, one of which is present in "sex-reversed" females. They suggest three sex chromosome constitutions in females of that species: XX, X*X, and X*Y. where X* represents a chromosome with a mutant short arm (X*p) that blocks testicular differentiation; E. W. Herbst, K. Fredga, F. Frank, H. Winking, A. Gropp, Chromosoma 69, 185 (1978). Remarkably X*p is shorter than Xp, see text and compare with Fig. 1.
4. S. Ohno, Sex Chromosomes and Sex-Linked Genes (Springer-Verlag, New York, 1967).
5. R. V. Short, Int. J. Androl. Suppl. 2, 21 (1978).
6. S. N. Ghosh, P. N. Shah, H. M. Gharpure, Nature (London) 276, 180 (1978).
7. J. L. Simpson, Disorders of Sexual Differentiation. Etiology and Clinical Delineation (Academic Press, New York, 1976), J. German J. L Simpson, R. S. K. Chaganti, R. L. Summitt, L. B. Reid, 1. R. Merkatz, Science 202, 53
8. For complete details of the case, see R. Bernstein, T. Jenkins, B. Dawson, J. Wagner, G. Dewald, G. C. Koo, S. S. Wachtel, J. Med. Genet., in press.
9. It may be argued that structural testis-determinants are situated on the X chromosome and activated by genes on the Y; see, for example, J. L. Hamerton, Nature (London) 219, 910 (1968) R. A. McFeely, W. C. D. Hare, J. D. Biggers Cytogenetics 6, 242 (1967). If H-Y structural genes are X-situated, then failure of H-Y synthesis in the present study could be due to a position effect resulting from duplication of a segment of Xp.
10. This work was supported in part by grants from the Rockefeller Foundation (RF-77042) and NIH (Al 11982, CA 08748, HD 00171, and HD 10065). We thank Dr. C. Dewald of the Mayo Clinic for the computer analysis.
Return To Molecular Biology Main Page
Return To Biology Dept. Main Page
![]()
![]()
© Copyright
2000 Department of Biology, Davidson College, Davidson, NC 28036
Send comments, questions, and suggestions to: macampbell@davidson.edu