Human Reproduction vol. 11 no.11 pp.2371-2375, 1996
OPINION
Valerie J.Grant
Department Psychiatry and Behavioural Science, School of Medicine, University of Auckland, New Zealand
The maternal dominance hypothesis has been derived from work with humans which shows that women who are more dominant than other women are more likely to conceive sons. In both animals and humans dominance is a characteristic or personality trait, underpinned by testosterone and responsive to a range of environmental changes: physical, social and psychological. Studies of the sex ratio in the social sciences and animal behaviour either support or are compatible with the idea that the sex-determining role of X- and Y- chromosome bearing spermatozoa may be preceded by factors under maternal control which provide for differential access of spermatozoa. Findings in reproductive physiology and physiological psychology suggest that follicular testosterone or a related hormone may play a critical role. Reproductive physiologists have already identified maternal mechanisms which could provide the context for such a model.
Key words: maternal dominance/sex determination/sex ratio
Introduction
In the population as a whole, the perception is that the sex of the infant is a matter of chance depending on whether an X or a Y chromosome bearing spermatozoon fertilizes the ovum. Furthermore, many researchers working on the determination of the sex of the infant appear to believe the same, since much of the work concentrates on the problems involved in identifying and separating X- and Y-chromosome-bearing spermatozoa (Beernink et at, 1993; Windsor et al., 1993; Ericsson, 1994). From this it might appear that reproductive physiologists are either unaware of parallel work being done in the social sciences, or not convinced of the relevance of the findings.
The unexplained male bias in the human secondary sex ratio (SSR) has provided the starting point for much of the work, and ensured that the determination of the sex of the infant continues to be a topic of almost universal interest. Demographers, anthropologists, psychologists and evolutionary biologists (among others) have sought to locate and describe unusual sex ratios in a variety of settings with a view to offering clues to reproductive physiologists as well as satisfying their own curiosity. Unlikely as it may seem, a synthesis of these findings at the present time points consistently in the direction of a role for the mother. When the current research emphasis in reproductive physiology points in one direction and that in the social sciences in the other, there is an opportunity to gain further insight from scrutinizing the competing claims.
One of the strongest of these arises from the maternal dominance hypothesis which was formulated on the basis of the results of a series of studies carried out over the last 30 years. The results provide consistent, statistically significant evidence to support the notion of a maternal involvement in the predetermination of the sex of her infant (Grant 1990, 1994a).
Measuring maternal dominance
The personality trait 'dominance' may be defined as influential,
ascendant, prevailing, authoritative, or high in control. It must
be discriminated from aggressive (hostile, angry, violent, quarrelsome,
argumentative) and domineering (overbearing, bossy, dictatorial
and high-handed) (Sadalla et al., 1988), neither of which is relevant.
Personality theorist Fiske (1971) defined the core of dominance
as 'acting overtly so as to change the views or actions of another'.
The personality tests, self report pencil and paper measures,
were constructed to measure the dominance of a particular individual
woman relative to other women (for further details of test theory
and construction see Grant, 1992).
Subjects were all healthy women (not seeking medical advice), and in studies undertaken after 1970, either < 8 weeks pregnant or pre-pregnant. This was to ensure that fetal testosterone in the maternal circulation, which becomes detectable by the 8th week of pregnancy (Edwards, 1980), was eliminated as an alternative interpretation of results. Because it is likely that the physiological basis of dominance is influenced by the social and psychological environment, pre-pregnant subjects were within 8 weeks of conception. Dates were calculated by working back from the date of birth of a normal full-term baby to the date on which the questionnaire was completed.
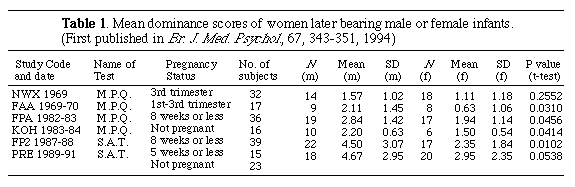
N (m) = No. of
women in the study who later gave birth to male infants.
N (f) = No. of women in the study who later gave birth
to female infants.
M.P.Q. = Dominance measured by means of the Maternal Personality
Questionnaire.
S.A.T. = Dominance measured by means of the Simple Adjective Test.
Mean (m) and SD (m) = Mean score and SD for women later bearing
male infants.
Mean (f) and SD (f) = Mean score and SD for women later bearing
female infants.
In six separate studies undertaken between 1969 and 1991, results were in the same direction and in all studies since 1970, at or near the same level of significance (P <0.05) (see Table I). A meta-analysis of all studies showed that women who later bore sons were significantly more likely to have scored higher on the tests of dominance than those who later bore daughters (X2 (12) = 89.5715, P <0.0001). When the two earliest studies (NWX, 1969, FAA, 1969-70), included in the table for the sake of completeness, are excluded because their pregnancies were advanced beyond the eighth week, the level of probability stays the same (X2 (8) = 64.6684, p <0.0001).
The maternal dominance hypothesis
The consistency and strength of these results led to the formulation
of the maternal dominance hypothesis, which states that women
who are more dominant in personality (when compared with the other
women in their cultural group) are more likely to conceive and
bear sons. The hypothesis is underpinned by two further propositions.
First, that dominance is a biologically-based personality trait
which humans share with animals (Wilson, 1975; Buss, 1988). Second,
that individual differences in dominance behaviour are highly
correlated with individual differences in concentrations of serum
testosterone. Although most of this research has been done with
male subjects, there are now several reports of the same phenomenon
in females (e.g. Purifoy and Koopmans, 1979; Baucom et al., 1985;
Udry and Talbert, 1988; Cashdan, 1995). This in turn is further
supported by the fact that intra-individual, environmentally-induced
changes in status-related dominance behaviours result in corresponding
changes in serum testosterone levels (e.g. Dabbs, 1988; Mazur,
1994). This means that both inter- and intra-personality differences
arise from a complex two way interaction between biological as
well as environmental influences (e.g. Udry et al., 1995), thus
providing the basis for change over time.
The maternal dominance hypothesis seeks to account for the fact that many women have infants of both sexes by postulating a normal distribution for dominance and its hormonal basis. It is known that environmental stimuli (social events) evoke changes in both dominance behaviours and the corresponding testosterone levels (again in both humans and animals; Rose et al., 1975; Mazur, 1976).
These factors mean that each individual woman would fluctuate in dominance, and the hormonal basis for dominance would fluctuate too. These fluctuations over time would be small to moderate in amount, as she responded to life events and other changes. For women scoring more than one standard deviation outside the normal distribution curve, there would be a tendency for these fluctuations not to alter the sex of consecutive infants. However for most women, whose scores lay within one SD, normal fluctuations in dominance (and testosterone) would take them either side of a critical threshold or amount which led to the conception of a male or female infant.
Compatibility with findings in reproductive physiology
This hypothesis implies a major maternal influence on the predetermination
of the sex of her infant. In the absence of knowledge of a clear
mechanism, it has been necessary, over the years, to monitor findings
in this area to ensure that the hypothesis was at least compatible
with findings in reproductive physiology. During the past three
decades a number of researchers have made suggestions which provided
a possible role for the female in determining the sex of her infant.
In general, these postulated differential access, under the control
of the mother for either X- or Y-chromosome-bearing spermatozoa.
Shettles' suggestions (1960, 1970) provided for both a male and a female role in the determination of the sex of the infant. Although accompanied by much publicity this work was later shown to be methodologically unsound (Gledhill, 1988). Reliable work includes hypothesized individual differences in the uterine environment, either chemical or mechanical or both (see for example Eisenbach and Tur-Kaspa, 1994); or maternal factors related to the timing of intercourse within the menstrual cycle, (see Gray, 1991 for a review of timing studies); or differences in the length of the follicular phase (Weinberg et al., 1995). The authors of this last paper described three possible ways in which their data could be explained, the first of which was that "the ovum produced in a cycle with a short follicular phase might be more receptive to Y- than to X-bearing spermatozoa, with the reverse being true in cycles with long follicular phases".
Saling (1991) also postulates maternal control of differential spermatozoan access through the zona pellucida, and Mittwoch (1989) suggests there may be a maternal factor regulating the rate of growth of the embryo in the immediate post-consception phase. The findings of both Mittwoch (1989) and Weinberg et al. (1995) support the data presented by Tarin et al. (1995) who found that "sex selection may be inadvertently performed in human in vitro fertilization (IVF) and embryo transfer programmes when selecting for high quality embryos (those with the fastest cleaving rates and/or the best morphology) at the fresh transfer cycle". In this work faster development was again associated with higher sex ratios. Any or all of the above findings could provide a physiological context which would increase the plausibility of the maternal dominance hypothesis.
Maternal testosterone
Although there is one study showing that maternal serum testosterone
varies in the predicted direction with the sex of the infant before
the eighth week of pregnancy (Harrison and Mansfield, 1980), there
do not appear to be any others. However, serum testosterone may
be important only as an indicator of follicular testosterone,
known to occur at a much greater concentration. According to Greenspan
and Baxter (1994) "The concentrations of luteinizing hormone
(LH) and follicle stimulating hormone (FSH) in follicular fluid
are generally lower than corresponding serum levels, but the concentrations
of ovarian steroids-androstenedione, testosterone, dihydrotestosterone
and estradiol-may be 10-40,000 times higher than their serum levels"
(p. 431). In describing androgen (and oestradiol) concentrations
in the late follicular phase of the menstrual cycle they say that
it is not known whether all these are part of development or play
"an important role in the process that leads to selection
of a single preovulatory follicle" (p. 431).
Haning et al. (1993) have demonstrated that follicular testosterone plays "a role previously unrecognized in the human as a major regulator controlling follicular aromatase" (p. 714). They also say that "these findings have important implications for understanding the control of follicular recruitment and atresia and the selection of the dominant follicle in humans" (p.710). If follicular testosterone or one of its related hormones has a role in determining which follicle becomes dominant and if Saling's (1991) suggestion about the ovum having control of spermatozoan entry through the zona pellucida is correct, then this provides a possible, plausible pathway towards suggesting that maternal factors may be playing a major role in the predetermination of the sex of the infant.
All the above findings refer to events before or at conception, suggesting a role for the mother in the predetermination of the sex of her infant by means of ovum selection and/or differential spermatozoan access. But this is not the only time when the mother could influence the sex ratio. Citing many animal studies Krackow (1995) argues first, that "the uterus is prepared by the dynamics of reproductive hormones to respond to implantation signals during a specific part of the oestrus cycle", secondly that "there is considerable variation in developmental pace between blastocysts" and thirdly, "the crucial point", that "the distribution of developmental stages may not be random with respect to the sex of blastocysts". Thus Krackow argues that shifts in the secondary sex ratio may be under maternal control at the implantation stage.
This idea is consistent with what is known about both early pregnancy loss and the likely primary sex ratio. In a prospective study Miller et al.. (1980) found a post-implantation pregnancy loss of 43% in a group of 197 normal women who had stopped contraception in order to conceive. It is also known that "Approximately 15% of registered pregnancies end in spontaneous abortion" (Nielsen and Hahlin, 1995). Amongst these losses it is highly likely that males are over-represented since the primary sex ratio is almost certainly higher than the secondary sex ratio. Hassold et al.. (1983) found in their study of 458 chromosomally normal spontaneous abortions, after allowing for contamination from maternal cells, that the primary sex ratio was likely to be in favour of males. Before the removal of hydatidiform moles the sex ratio was 108, and after their removal, 116, compared with the normal secondary sex ratio of 105 or 106 males for every 100 females.
Thus Krackow's "developmental asynchrony hypothesis for sex ratio manipulation" is at least plausible in terms of the wider parameters. But there would seem no need to choose between this and the maternal dominance hypothesis since the latter is said to be relevant before or at conception (i.e. it postulates the predetermination of the sex of the infant (male) by means of an ovum specially designed to receive a Y chromosome-bearing spermatozoon or nothing), and the former which postulates maternal mechanisms which operate to differentially foster XX or XY embryos after fertilization. It seems reasonable to admit both hypotheses, the one operating before and the after conception.
Given the consistency with which dominance behaviours have been found to correlate with the conception of male infants; the prominent role that testosterone appears to take both in providing the physiological basis for dominance behaviours, and possibly also in the selection and development of a particular ovum; and the possibility that different developmental rates may be synchronized with conditions in the maternal reproductive tract, further investigation of the role of the mother in the predetermination of the sex of her infant and its continuing development would seem to be justified.
Evidence from other sources
Supportive evidence for the maternal dominance hypothesis comes
from a number of other social sciences. Either the findings have
already suggested a role for the female or a reinterpretation
of the data in terms of the maternal dominance hypothesis is as
plausible as the original. The most important of these areas are
the studies of the human SSR (see for example Stern, 1973). Given
that there is no evidence to suggest that X- and Y-chromosome-bearing
spermatozoa are produced in anything except equal numbers, why
would the SSR fall consistently between 105:100 and 106:100 in
favour of males? If the SSR had been the same in humans as it
is in other mammals (100:100) it would probably not have attracted
so much attention.
Much early work was directed at discovering whether or not there is a genetic influence on the sex ratio. Investigations were carried out into the occurrence of single-sex sibships and family tendency to produce either male or female infants in successive generations. No evidence for a genetic effect was found (Edwards, 1962; Teitelbaum, 1972). Tiny variations, discernible only on population sample sizes of several millions are those connected with race, parity, and socioeconomic status (Chahnazarian, 1988; James, 1987).
However, in 1985 James made a theoretical prediction that dominance and sex ratios could be related. By comparing findings across disciplines (demography and evolutionary biology) he suggested that sex ratios could be associated with dominance rank in some primates since both could be associated with hormone levels at the time of conception. This observation allowed for the re-emergence of the question about a genetic influence on the sex ratio. It was hinted at again the following year by Clutton-Brock and Iason (1986) when they sought to explain known variations in animal and human sex ratio data. "In view of the apparent lack of genetic variance in the sex ratio in many species" they wrote, "a hormonal mechanism mediated by environmental factors provides a plausible explanation of many trends".
Since that time there have been many papers documenting a relationship between dominance rank and altered sex ratios in animals (for example, Clutton-Brock et al., 1984; Meikle et al., 1984; Aureli et al., 1990; Meikle et al., 1993). Many of the papers present data from longitudinal studies carried out over periods of several years. They present statistically significant findings on the sex ratio of offspring according to dominance rank of the mother. On the other hand, results have been conflicting and are said by some (Smith and Smith, 1985) to be indicative only of major sampling error. These studies are now being re-analysed and reinterpreted on the basis of the hypothesized proximate mechanisms described above (Grant, unpublished).
One small, specialist part of the work with animals and sex ratio is not disputed. It is known that the intrauterine position of a rodent female fetus has an influence on the sex of her offspring. "Female fetuses that mature between males are androgenized by testosterone crossing fetal membranes". When mature these animals "produce litters containing a significantly greater proportion of sons than the litters produced by those that gestated between female fetuses" (Clark et al., 1993). The effect has been found independently by other researchers in gerbils and house mice (Vandenburgh and Huggett, 1994). It offers the possibility of a genetic effect on the sex ratio (i.e. females might inherit a tendency towards higher or lower testosterone levels) as well as a clue about potential mechanisms. Like so many studies in this area, testosterone is not only the focus of interest but very small variations in the amount of testosterone appear to make the critical difference.
Thus the traditional "coin-tossing" model for the allocation of the sex of the infant, never entirely satisfactory because of the distorted human sex ratio, appears less and less satisfactory. Viewing the problem from the point of view of evolutionary biology it now appears far more likely that females conceive infants of the sex they are most suited to raise. In animals, aspects of the environment (e.g. temperature, see for example Haig, 1991); or local resource distribution (see for example Verme, 1969; Mech, 1975), as well as good physiological condition (Trivers and Willard, 1973) may be relevant. In humans, and possibly in other primates and ungulates, maternal dominance is likely to be an important factor.
The maternal dominance hypothesis is compatible with the major findings in the area of sex ratio; both those that reflect stability across large population samples and those which find variation in well defined small groups. The apparent lack of a genetic effect in the findings so far might be explained in future by re-working population models of sex ratio to show how the opposing effects of Poisson and Lexis variation have, after all, masked an underlying genetic effect (James, 1996).
A further area of support for the maternal dominance hypothesis comes from work in developmental psychology. In a corollary to the main hypothesis it is suggested that the personality of the mother is not only related to the sex of her infant through the hormones of reproduction, but is appropriate to the psychological development of the infant. If dominant mothers conceive and bear male infants, then one might postulate that male infants as a group were being exposed to mothers who, at least temporarily were different in their behaviour to that of mothers of female infants.
There is good, independent evidence to support this. In the search for the origins of psychological sex differences, developmental psychologists worked their way back to the beginning. Several different teams of researchers, from different countries and at different times all found the same thing; namely that mothers behave differently towards their babies according to the sex of the infant, and in the absence of differences in the babies' behaviours (see for example Moss, 1967; Hwang, 1978; Gottfried et al.., 1987). At the time theorists concluded that these differences were indicative of the pervasiveness and strength of sex stereotyping. Of course parental expectations will be playing a part, but there is also evidence that mothers were behaving in ways that were natural to them. The very nature of the differences (mothers of male infants were more initiating; mothers of female infants more responsive) were exactly what one would have predicted from the maternal dominance hypothesis. (For a full account see Grant, 1994b.)
Conclusions
Although reproductive physiologists appear to be emphasizing the role of spermatozoa in their work on the determination of the sex of the infant, it is likely that maternal factors are at least as important. Evidence to support a maternal dominance hypothesis comes primarily from results of human studies showing a relationship between the personality trait dominance and an increased likelihood of conceiving a male infant. The same effect has been observed in animal studies, although as yet the results are equivocal. In addition large scale population studies of the human secondary sex ratio, as well as some small scale animal studies, support or are compatible with the idea of maternal control of the sex ratio.
Work by physiological psychologists and animal behaviourists has demonstrated that testosterone provides a biological underlay for dominance in both animals and humans. Reproductive physiologists have described maternal mechanisms in the reproductive process which could be regulated by testosterone, thus providing a means by which spermatozoan access could be under maternal control.
Acknowledgement
I gratefully acknowledge the advice and assistance of Dr Lois Salamonsen, Prince Henry's Institute of Medical Research, Monash University, Melbourne, Australia.
References
Aureli, E, Schino, G., Cordischi, C. et al.. (1990) Social factors affect secondary sex ratio in captive Japanese Macaques. Folia Primatol., 55, 176-180.
Baucom, D.H., Besch, RK. and Callahan, S. (1985) Relation between testosterone concentration, sex role identity, and personality among females. J. Pers. Soc. Psych., 48, 1218-1226.
Beemink, EJ., Dmowski, W.R and Ericsson R.J. (1993) Sex preselection through albumin separation of sperm. Fertii Steril., 59, 382-386.
Buss, A.H. (1988) Personality: Evolutionary Heritage and Human Distinctiveness Erlbaum, Hillsdale, NJ, USA.
Cashdan, E. (1995) Hommones, sex and status in women. Horm. Behav., 29 354-366.
Chahnazarian, A. (1988) Detemminants of the sex ratio at birth: Review of recent literature. Soc. Biol., 35, 214-235.
Clark, M.M., Karpluk, R and Galef, B.G. (1993) Hommonally mediated inheritance of acquired characteristics in Mongolian gerbils. Nature, 364, 712.
Clutton-Brock, T.H. and lason, G.R. (1986) Sex ratio variation in mammals. QuarL Rev. Biol., 61, 339-374.
Clutton-Brock, T.H., Albon, S.D. and Guinness, F.E. (1984) Matemal dominance, breeding success and binh sex ratios in red deer. Nature, 308, 358-360.
Dabbs, J.M., Ruback, R.B., Hopper, C.H. and Sgoutas, D.S. (1988) Saliva testosterone and criminal violence among women. Pers. Ind. Diffs., 9, 269-275.
Edwards, A.W.E (1962) Genetics and the human sex ratio. Adv. Cenet., Il. 239-272.
Edwards, R.G. (1980) Conception in the Human Female. Academic Press, London.
Eisenbach, M. and Tur-Kaspa, I. (1994) Human spemm chemotaxis is not enigmatic anymore. Fertil. Steril., 62, 233-235.
Ericsson, R.J. (1994) Sex selection via albumin columns: 20 years of results. Hum. Reprod., 9, 1787-1791.
Fiske, D.W. (1971) Measuring the Concepts of Human Personality. Aldine, Chicago.
Gledhill, B.L. (1988) Selection and separation of X- and Y-chromosome bearing mammalian spemm. Gamete Res., 20, 377-395.
Gottfried, N.W., Seay, B.M. and Wismar, K. (1987) Neonatal sex and binhorder effects on behaviours of mothers and temporary caretakers during bottle-feeding. J. Genet. Psych., 148, 479-487.
Grant, V.J. (1990) Matemal personality and sex of infant. Br J. Med. Psych., 63, 261-266.
Grant, V.J. (1992) The measurement of dominance in preonant women by use of the Simple Adjective Test. Person. Individ Diff., 13, 99-102.
Grant, V.J. (1994a) Matemal dominance and the conception of sons. Br J. Med. Psych., 67, 343-351.
Grant, V.J. (1994b) Sex-of-infant differences in mother-infant interaction: a reinterpretation of past findings. Dev. Review, 14, 1-26.
Gray, R.H. (1991) Natural family planning and sex selection: fact or fiction? Am. J. Obstet. Gynecol., 165, 1982-1984.
Greenspan, ES. and Baxter, J.D. (eds) (1994) BasEc and Clinical Endocrinology. 4th edn. Prentice-Hall Int., London.
Haig, D. (1991) Developmental asynchrony and environmental sex detemmination in alligators. J. Theor Biol., 150, 373-383.
Haning, R.V., Hackett, R.J., Elood, C.A. et al.. (1993) Testosterone: a follicular regulator: key to anovulation. J. Clin Endocrinol. Metab., 77, 710 715.
Harrison, R.E and Mansfield, M.D. (1980) Matemal plasma androgens in early human pregnancy. Br. J. ObsteL Gynaecol., 87, 695-704.
Hassold, T., Quillen, S.D. and Yamane, J.A. (1983) Sex ratio in spontaneous abonions. Ann. Hum. Genet., 47, 39-47.
Hwang, C.-P. (1978) Mother-infant interaction: Effects of sex of infant on feeding behaviour. Early Hum. Dev., 2, 341-349.
James, W.H. (1985) Sex ratio, dominance status and matemal hommone levels at the time of conception. J. Theor Biol., 114, 505-510.
James, W.H. (1987) The human sex ratio 1: a review of the literature. Hum. Biol., 59, 721-752.
James, W.H. (1996) Evidence that mammalian sex ratios at binh are panially controlled by parental hommone levels at the time of conception. J Theor. Biol., 271-286.
Krackow, S. (1995) The developmental asynchrony hypothesis for sex ratio manipulation. J. Theor. Biol., 176, 273-280.
Mazur, A. (1976) Effects of testosterone on status in primate groups. Folia Primatol., 26, 214-226.
Mazur, A. (1994) A neurohormonal model of social stratification among humans: a microsocial perspective. In Ellis, L. (ed.), Social Stratification and Socioeconomic Inequality. Vol 2. Reproductive and Interpersonal Aspects of Dominance and Status. Praeger, Westpon, Connecticut.
Mech, L.D. (1975) Disproponionate sex ratios in wolf pups. J. Wildlife Manag., 39, 737-740.
Meikle, D.B, Tflford, B.L. and Vessey, S.H. (1984) Dominance rank, secondary sex ratio and reproduction of offspring in polygynous primates. Am. Nat, 124, 173-188.
Meikle, D.B., Drickamer, L.C., Vessey, S.H. et al.. (1993) Maternal dominance rank and secondary sex ratio in domestic swine. Anim. Behav., 46, 79-85.
Miller, J.E, Williamson, E. Glue, J. et al.. (1980) Fetal loss after implantation. Lancet, ii, 554-556.
Mittwoch, U. (1989) Sex differentiation in mammals and tempo of growth: probabilities vs. switches. J. Theor Biol., 137, 445-455.
Moss, H.A: (1967) Sex, age, and state as detemminants of mother-infant interaction. Merr-Palmer QuarL, 13, 19-36.
Nielsen, S. and Hahlin, M. (1995) Expectant management of first-trimester spontaneous abonion. Lancet, 345, 84-86.
Purifoy, EE. and Koopmans, L.H. (1979) Androstenedione, testosterone, and free testosterone concentration in women in various occupations. Soc. Biol., 26, 179-188.
Rose, R.M., Bemstein, I.S. and Gordon, T.R (1975) Consequences of social conflict on plasma testosterone levels in rhesus monkeys. Psvehosom. Med. 37, 50-60.
Sadalla, E.K., Kenrick, D.T. and Vershure, B. (1987) Dominance and heterosexual attraction. J. Pers. Soc. Psych., 52, 730-738.
Saling, RM. (1991) How the egg regulates spemm function dunng gamete interaction: facts and fantasies. Biol. Reprod, 44, 246-251.
Shenles, L.B. (1960) Nuclear morphology of human spemmatozoa. NatuK, 186, 648-649.
Shettles, L.B. (1970) Factors influencing sex ratios. InL J. Gynecol. Obstet., 8, 643-647.
Smith, M.E and Smith, D.G. (1985) Sex of infants produced by male rhesus macaques. Am. Nat., 126, 354 361.
Stem, C. (1973) The sex ratio. In Principles of Human Genetics. Freeman, San Francisco, Chapter 21.
Tarin, J.J., Bemabeu, R., Baviera, A. et al.. (1995) Sex selection may be inadvenently performed in in-vitro fertilization-embryo transfer programmes. Hum. Reprod, 10, 2992-2998.
Teitelbaum, M.S. (1972) Factors associated with the sex ratio in human populations. In Harrison, G.A. and Boyce, A.J. (eds), The Structure of Human Populations. Clarendon Press, Oxford.
Trivers, R.L. and Willard, D.E. (1973) Natural selection of parental abiiity to vary the sex ratio of offspring. Science, 179, 90-91.
Udry, J.R. and Talben, L.M. (1988) Sex hommone effects on personality at pubeny. J. Pers. Soc. Psych., 54, 291-295.
Udry, J.R., Morris, N.M. and Kovenock, J. (1995) Androgen effects on women's gendered behaviour. J. Biosoc. Sci., 27, 359-368.
Vandenbergh, J.C. and Huggett, C.L. (1994) Mother's prior intrauterine position affects the sex ratio of her offspring in house mice. Proc. Natl. Acad Sci USA, 91, 11055-11059.
Vemme, L.J. (1969) Reproduction patems of white-tailed deer related to nutritional plane. J. Wildlife Manag., 33, 881-887.
Weinberg, C.R., Baird, D.D. and Wilcox, A.J. (1995) The sex of the baby may be related to the length of the follicular phase in the conception cycle. Hum. Reprod, 10, 304-307.
Wilson, E.O. (1975) Sociobiology: The New Synthesis. Harvard University Press, Cambridge, USA.
Windsor, D.R, Evans, G. and White, I.G. (1993) Sex predetemmination by separation of X and Y chromosome-bearing sperm: a review. Reprod Fertil. Dev., 5, 155-171.
Received on April 9, 1996; accepted on September 11, 1996
Return To Molecular Biology Main Page
Return To Biology Dept. Main Page
![]()
©
Copyright 2000 Department of Biology, Davidson College, Davidson,
NC 28036
Send comments, questions, and suggestions to: macampbell@davidson.edu