Gene, 111 (1992) 229-233
©1992 Elsevier Science Publishers B.V. All rights reserved. 0378-1119/92/$05.00
GENE 06296
SUMMARY
Many cnidarians utilize green-fluorescent proteins (GFPs) as energy-transfer
acceptors in bioluminescence. GFPs fluoresce in vivo upon receiving
energy from either a luciferase-oxyluciferin excited-state complex or a
Ca2+-activated photoprotein.
These highly fluorescent proteins are unique due to the chemical nature
of their chromophore, which is comprised of modified amino acid (aa) residues
within the polypeptide. This report describes the cloning and sequencing
of both cDNA and genomic clones of GFP from the cnidarian, Aequorea victoria.
The gfp10 cDNA encodes a 238-aa-residue polypeptide with a calculated
Mr of 26, 888. Comparison of A. victoria
GFP genomic clones shows three different restriction enzyme patterns which
suggests that at least three different genes are present in the A. victoria
population at Friday Harbor, Washington. The gfp gene encoded by the l
GFP2 genomic clone is comprised of at least three exons spread over
2.6 kb. The nucleotide sequences of the cDNA and the gene will aid in the
elucidation of structure-function relationships in this unique class of
proteins.
INTRODUCTION
Luminescence is common in a variety of marine invertebrates. Many cnidarians
and probably all ctenophores emit light when mechanically disturbed. Proteins
responsible for bioluminescence from several species of these two phyla
have been characterized. Light from luminescent cnidaria is primarily green
whereas light emitted from ctenophores is blue. The green light of cnidaria
is due to the presence of a class of proteins called green-fluorescent proteins
(GFPs). They are highly fluorescent and are activated in vivo by
an energy transfer process via a luciferase or a Ca2+
-activated photoprotein, both of which produce energy during the oxidation
of coelenterate-type luciferin. In the cnidarian Aequorea, the photoprotein
aequorin excites the GFP by an unknown mechanism to release green light.
Previous studies suggesting that Aequorea GFP is stimulated via a
radiationless mechanism (Morise et al., 1974) have been questioned (Ward,
1979). The GFP from Renilla, another cnidarian, on the other hand,
clearly receives energy from the Renilla luciferase-oxyluciferin
excited state complex by a radiationless energy transfer mechanism (Ward
and Cormier, 1976).
The GFPs most thoroughly studied have been isolated from Aequorea
and Renilla (Ward, 1979). The Aequorea GFP has been reported
to be a 30-kDa monomer (Prendergast and Mann, 1978) whereas the Renilla
GFP is a 54-kDa homodimer (Ward and Cormier, 1979). The two proteins have
different absorption spectra but identical emission spectra (l
max = 509 nm). Upon denaturation, the two GFPs have the same absorption
spectra. Ward et al. (1980) have predicted that both Aequorea and
Renilla GFPs contain chromophores having the same structure but that
the different absorption spectra are explained by different apoprotein environments.
Biochemical properties of the Aequorea GFP show it to have unique
structural properties. The fluorescent chromophore is stable to a variety
of harsh conditions including heat, extreme pH, and chemical denaturants.
Fluorescence is lost, for example, to base or acid treatment or addition
of guanidine hydrochloride, but upon neutralization of the pH or removal
of the denaturant, fluorescence returns with an identical emission spectrum
(Bokman and Ward, 1981; Ward and Bokman, 1982). The chromophore structure
is very different from those of the phycobiliproteins which are also highly
fluorescent. The chromophore in the GFPs is covalently bound and is formed
by modification of certain aa residues within the polypeptide. The chemical
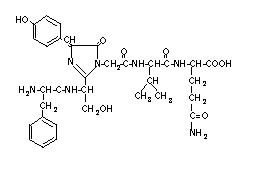
structure of the Aequorea GFP chromophore (Fig. 1), first characterized
by Shimomura (1979), has been thoroughly re-examined (Ward et al.,
1989; W.W.W., unpublished) and is shown here (Fig. 1) in its revised form.
In this study, the Aequorea GFP gene and its cDNA have been isolated
and characterized in pursuit of elucidating the mechanism of energy transfer
between aequorin and GFP as well as addressing evolutionary relationships
in coelenterate bioluminescence.
Fig. 1. The chemical structure of the chromophore in Aequorea GFP (W.W.W., unpublished). The cyclized chromophore is formed from the trimer Ser-dehydroTyr-Gly within the polypeptide by an unknown mechanism.
EXPERIMENTAL AND DISCUSSION
(a) Construction of cDNA libraries
An A. victoria cDNA library, constructed in pBR322 (Prasher et al.,
1985), was screened for the presence of a gfp cDNA using two oligo mixtures
whose sequences were based on the aa sequences derived from GFP-derived
CNBr fragments. The oligos contained the following nt sequences: A: 5 '
![]() (20-mer with 32 redundancies), B: 5 '
(20-mer with 32 redundancies), B: 5 ' ![]() (17-mer with
16 redundancies). The hybridization of the 32P-labeled
mixtures A and B to replicate filters containing this library were performed
according to the method of Wood et al. (1985) utilizing tetramethylammonium
chloride during the washing steps. The temperatures used during the washing
steps for mixtures A and B were 55 ° C and 50° C, respectively.
(17-mer with
16 redundancies). The hybridization of the 32P-labeled
mixtures A and B to replicate filters containing this library were performed
according to the method of Wood et al. (1985) utilizing tetramethylammonium
chloride during the washing steps. The temperatures used during the washing
steps for mixtures A and B were 55 ° C and 50° C, respectively.
A single gfp cDNA was isolated from the library by this method. This clone,
pGFP1, contained a PstI insert of 511bp having an ORF encoding 168 aa. The
deduced translation of the nt sequence indicated the gfp1 cDNA lacked
both the 5 ' - and 3' -sequences of the coding region. However, the sequence
FSYGVQ within the deduced translation permitted the chromophore structure
to be deciphered (W.W.W., unpublished). Upon rescreening the library with
gfp1 cDNA, no additional cDNAs were found.
A second A . victoria cDNA library was constructed (Gubler and Hoffman,
1983) in l gt10 (Huynh et al., 1985). The PstI
insert from gfp1 cDNA was used as a hybridization probe against the
entire l gt10 library of 1.4 x 106
recombinant phage. No gfp-related recombinants were identified upon
screening the primary library. The phage remaining on the plates were extracted
from the top agar and used as an amplified library (Maniatis et al., 1982).
Upon screening this preparation of the library, four recombinants hybridized
to the gfp1 cDNA following their purification. The four cDNA clones
were designated ![]() GFP10, 11, 12. and 13. All four recombinants were shown
to contain an insert of 1 kb upon digestion with EcoRI.
GFP10, 11, 12. and 13. All four recombinants were shown
to contain an insert of 1 kb upon digestion with EcoRI.

Fig. 2. Nucleotide sequence of the gfp10 cDNA and the deduced aa sequence. Below the first nt of each codon is the single-letter designation for the aa. The horizontal lines underline those aa sequenced directly from native GFP. The downward arrows indicate the positions of introns when compared to the nt sequence of the gfp2 gene. Arrowhead: start codon; period: stop codon. DNA fragments from both cDNA and genomic clones were subcloned into M13mp18 and M13mp19 (Yanisch-Perron et al., 1985), and unidirectional deletions were prepared using the method of Dale et al. (1985). Sequencing was performed using either the Klenow fragment or an altered T7 DNA polymerase (Sequenase Ver 2.0, United States Biochemical Corp.) in the dideoxy chain termination method (Sanger et al., 1977). Both DNA strands of the sequences described in this report here have been sequenced. The GenBank accession No. for the gfp10 sequence is M62653.
(b) Characterization of the gfp10 cDNA
The entire EcoRI insert of l GFP10 was sequenced
(Fig. 2). Limited nt sequences obtained from l GFP
11 and 12 were identical with that from l GFP10
suggesting that they were siblings and, hence, were not sequenced further.
Even though the entire coding region appears to be present (see below),
three features of the cDNA insert of l GFP10
suggest it is not quite full-length. First, the cDNA is 965 nt where the
gfp mRNA is 1.05 kb in length as determined by Northern analysis
(Fig. 3). Second, the 5'-untranslated region is very short. Third, no poly(A)+
track is observed in the gfp10 cDNA sequence (Fig. 2) despite the
presence of the gfp mRNA in only the poly(A)+ RNA
fraction of A. victoria RNA (data not shown). A typical polyadenylation
signal is located at nt 861-865 (Fig. 2).
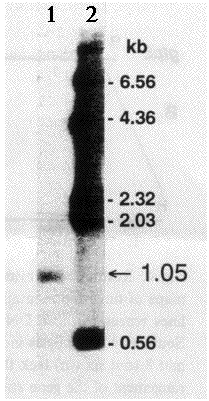
Fig. 3. Northern analysis of the A. victoria gfp mRNA. The poly(A)+ mRNA (lane 1) was denatured using glyoxal prior to electrophoresis, as described by Thomas (1983). Electrophoresis was performed for 3 h in a 1 % agarose gel (pretreated with 10 mM sodium phosphate) equilibrated in 10 mM sodium phosphate pH 7.0 buffer. Overnight transfer of the nucleic acids to nitrocellulose was facilitated with 20X SSC. Hybridization of 32P-labeled gfp1 cDNA to the membrane-bound nucleic acids was at 42° C for 28 h in 5X SSC/5X Denhardt's/20 mM Na.phosphate pH 6.8/100 µg per ml of denatured herring sperm DNA/10% polyethyleneglycol/50% formamide. HindIII-digested l DNA, 32P-labeled, and treated in parallel with the RNA, was used as molecular weight standards (lane 2).
The nt sequence of the gfp10 cDNA contains an ORF encoding a 238-aa
protein having a calculated Mr of 26,888.
This compares favorably with 30 kDa for native GFP as determined by denaturing
electrophoresis (Prendergast and Mann, 1978). The deduced translation contains
aa sequences of numerous peptides isolated from native GFP (underlined in
Fig. 2). When compared to the gfp10 cDNA sequence (Fig. 2), the gfp1
cDNA was determined to encode aa residues 28-195. Oligo mixture A is complimentary
to the codons encoding aa 78-84 and mixture B is complimentary to the codons
encoding aa 141-146 (Fig. 2). The trimer Ser-Tyr-Gly, modified in the native
protein to form the chromophore (W.W.W., unpublished), is located at aa
65-67. The chromophore consists of an imidazolone ring formed by the residues
Ser-dehydroTyr-Gly within the polypeptide (Fig. 1). Located 8 aa upstream
of this chromopeptide is GFP's only Trp. The inability to detect the fluorescence
from this Trp makes it unusual (W.W.W., unpublished). Perhaps energy-transfer
occurs between it and the chromophore in the native protein preventing the
Trp fluorescence (320-350 nm). The Trp is flanked by several Pro residues
(Pro-Val-Pro-Trp-Pro). The significance of this pentapeptide is not understood
but a search of the protein databases (PIR ver 25; Swiss-Prot ver 14) shows
it to be present only in cytochrome P-450 proteins.
(c) Isolation and characterization of gfp genomic clones
The gfp1 cDNA was also used to isolate genomic clones prior to the
availability of the gfp10 cDNA. An A. victoria genomic library
was constructed in l 2001 (Karn et al., 1984)
essentially as described (Maniatis et al., 1982). Eight recombinant phages
hybridizing to the gfp1 cDNA were purified from the genomic DNA library.
Based on restriction enzyme and Southern-blot analyses, they represent six
different isolates having at least three different restriction maps (Fig.
4). When DNA fragments from the 5'- and 3'-ends of the gfp1 cDNA
were used as hybridization probes, all of the genomic clones were found
likely to contain the 5'-end of the gene, but only gfp2, 3, and 9
also contained the 3 ' end. The three types of genomic clones are consistent
with the presence of multiple GFP isoforms isolated from A. victoria
(A. Roth, M. Cutler and W.W.W., unpublished). Since the A. victoria
genomic DNA used for the genomic library was isolated from a large number
of jellyfish (collected at Friday Harbor, Washington), the three gfp
genes are representative of the Aequorea population as opposed to
individual jellyfish.
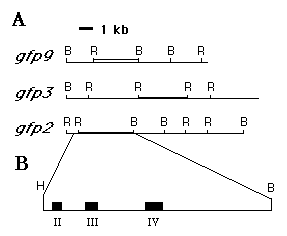
Fig. 4. Restriction enzyme maps of three Aequorea gfp genes. (A) The maps of three representative genomic clones are compared. The double lines represent those DNA fragments which hybridize to gfp1 cDNA. Southern-blot analysis indicated three other genomic clones, l GFP1, 4 and 8 (not shown) lack the 3' end of the gene. (B) The exon/intron arrangement of the gene encoded by l gfp2 was determined by comparing the nt sequences of the 5-kb EcoRI - BamHI and the overlapping 1.8-kb HindIII fragments of l GFP2 and the EcoRI insert of l GFP10 cDNA. The exons are represented by the blackened boxes, II, III, and IV. The GenBank accession No. for the gfp2 sequence is M62653.
The EcoRI-BamHI and an overlapping HindIII fragments in the genomic clone l GFP2 (Fig. 4) were sequenced and compared to that of the gfp10 cDNA to examine the structure of the gene. The gfp gene encoded by l GFP2 contains at least three exons spread over 2.6 kb of DNA (Fig. 4). These exons, designated II, III, and IV, encode 69, 98, and 71 aa, respectively. Presumably, a fourth exon is located upstream from the genome since the 15 nt at the 5' end of the gfp10 cDNA sequence cannot be aligned to the 5 ' region of the DNA sequence derived from the gfp2 gene. The positions of the introns with respect to the cDNA sequence are indicated (Fig. 2). The aa residues involved in the chromophore are encoded at the 3' end of exon II. The nt sequences of the gfp mRNA splice junctions agree reasonably well with consensus sequences (Fig. 5).

Fig. 5. Alignment of the nt sequences in gfp2 at the splice junctions. The intron sequences were identified by comparing the nt sequences of gfp2 and the gfp2 cDNA (Fig. 2). The consensus sequence is taken from Senapathy et al. (1990).
The gfp10 cDNA is not encoded by the gfp2 gene since there are several nt differences between their sequences. The nt differences within the protein-coding regions are summarized in Table IA. Four of the 12 single nt differences result in conservative aa replacements at positions 100, 108, 141 and 219 (Table IB). The aa residues encoded at these four positions are consistent with the aa sequences observed in GFP-derived peptides which showed a Tyr at position 100, a Met at position 141, but a Thr at position 108. Eight additional nt differences occur with the gfp2 gene in the 3'-non-translated region of the gfp10 cDNA (data not shown). It is not known whether the gfp10 cDNA represents an allele of gfp2 or another gfp gene.
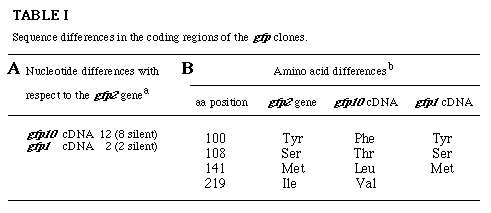
aTotal number observed upon comparison of
the nt sequencs of the ORFs in the gfp cDNAs with the homologous
sequencs in the gfp2 gene.
bObserved upon comparison of the translations of the ORFs
of both cDNAs and the exons of the gfp2 gene. The aa numbering is
the same as that used in Fig. 2.
These results will enable us to construct an expression vector for the
preparation of non-fluorescent apoGFP. Since no information is yet available
regarding the biosynthesis of the chromophore, a recombinant form of this
protein will be a valuable reagent with which to examine the biochemistry
of chromophore formation in this unique class of proteins and the mechanism
of energy transfer between aequorin and GFP.
ACKNOWLEDGEMENTS
We want to extend our thanks to Bonnie Woodward, Darlene Bianca, and Richard
McCann for their excellent technical assistance. Supported in part by a
Mellon Award from the Woods Hole Oceanographic Institution (27/50.44) and
a grant from the American Cancer Society (NP640) to D.C.P. A paper of the
journal series New Jersey Agricultural Experiment Station. This work was
performed as part of NJAES Project No. 01102. A special thanks goes to Dr.
A.O.D. Willows, Director, Friday Harbor Laboratories, for use of laboratory
facilities.
Correspondence to: Dr. D.C. Prasher, Redfield Bldg., Woods
Hole Oceanographic Institution, Woods Hole, MA 02543 (U.S.A.) Tel. (508)457-2000,
ext. 2311; Fax (508)457-2195.
Abbreviations: A., Aequorea; aa, amino acid(s); bp, base pair(s); GFP, green-fluorescent protein;
gfp, DNA or RNA encoding GFP; kb, kilobase(s) or 1000 bp; nt, nucleotide(s);
oligo, oligodeoxyribonucleotide; ORF, open reading frame(s).
REFERENCES
Bokman, S.H. and Ward, W.W.: Renaturation of Aequorea green fluorescent
protein. Biochem. Biophys. Res. Commun. 101 (1981) 1372-1380.
Dale, R.M.K., MeClure, B.A. and Houchins, J.P.: A rapid single-stranded
cloning strategy for producing a sequential series of overlapping clones
for use in DNA sequeneing: applieation to sequeneing the eorn mitochondrial
18S rDNA. Plasmid 13 (1985) 31-40.
Gubler, U. and Hoffman, BJ.: A simple and very efficient method for Denerating
cDNA libraries. Gene 25 (1983) 263-269.
Huynh, T.V., Young, R.A. and Davis, R.W.: Constructing and screening cDNA
libraries in l gt10 and l
gt11. In: Glover, D.M. (Ed.), DNA Cloning: A Practical Approach,
Vol. 1. IRL Press, Oxford, 1985, pp. 49-78.
Karn, J., Matthes, H.W.D., Gait, M.J. and Brenner, S.: A new selective phage
cloning vector, l 2001, with sites for XbaI,
BamHI, HindIII, EcoRI, SstI and XhoI. Gene 32 (1984) 217-224.
Maniatis, T., Fritsch, E.F. and Sambrook, J.: Molecular Cloning. A Laboratory
Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1982.
Morise, J.G., Shimomura, O., Johnson, F.H. and Winant, J.: Intermolecular
energy transfer in the bioluminescent system of Aequorea. Biochemistry
13 (1974) 2656-2662.
Prasher, D., McCann, R.O. and Cormier, M.J.: Cloning and expression of the
cDNA coding for aequorin, a bioluminescent calcium-activated protein. Bioehem.
Biophys. Res. Commun. 126 (1985) 1259-1268.
Prendergast, F.G. and Mann, K.G.: Chemical and physical properties of aequorin
and the green-fluorescent protein isolated from Aequorea forskalea.
Biochemistry 17 (1978) 3448-3453.
Sanger, F., Nicklen, S. and Coulson, A.R.: DNA sequencing with chain-terminating
inhibitors. Proc. Natl. Acad. Sci. USA 74 (1977) 54635467.
Senapafhy, P., Shapiro, M.B. and Harris, N.L.: Splice junctions, branch
point site, and exons: sequence statistics, identification, and applications
to genome project. Methods Enzymol. 183 (1990) 252-278.
Shimomura, O.: Structure of the chromophore of Aequorea green fluorescent
protein. FEBS Lett. 104 (1979) 220-222.
Thomas, P.S.: Hybridization of denatured RNA transferred or dotted to nitrocellulose
paper. Methods Enzymol. 100B (1983) 255-266.
Ward, W.W.: Energy transfer processes in bioluminescence. Photochem. Photobiol.
Rev. 4 (1979) 1-57.
Ward, W.W. and Bokman, S.H.: Reversible denaturation of Aequorea
green-fluorescent protein: physical separation and characterization of the
renatured protein. Biochemistry 21 (1982) 4535-4550.
Ward, W.W. and Cormier, M.J.: In vitro energy transfer in Renilla
bioluminescence. J. Phys. Chem. 80 (1976) 2289-2291.
Ward, W.W. and Cormier, M.J.: An energy transfer protein in coelenterate
bioluminescence. J. Biol. Chem. 254 (1979) 781-788.
Ward, W.W., Cody, C.W., Hart, R.C. and Cormier, M.J.: Spectrophotometric
identity of the energy-transfer ehromophores in Renilla and Aequorca
green-fluoreseent proteins. Photochem. Photobiol. 31 (1980) 611-615.
Ward, W.W., Cody, C.W., Prasher, D.C. and Prendergast, F.G.: Sequenee of
the chemical structure of the hexapeptide chromophore of Aequorea
green-fluorescent protein. Photochem. Photobiol. 49 (1989) 62S.
Wood, W.I., Gitschier, J., Lasky, L.A. and Lawn, R.M.: Base composition-independent
hybridization in tetramethylammonium chloride: a method for oligonucleotide
screening of highly complex gene libraries. Proc. Natl. Acad. Sci. USA 82
(1985) 1585-1588.
Yanisch-Perron, C., Vieira, J. and Messing, J.: Improved M13 phage cloning
vectors and host strains: nucleotide sequences of the M13mpl8 and pUCI9
vectors. Gene 33 (1985) 103-119.
![]()
© Copyright
2000 Department of Biology, Davidson College, Davidson, NC 28036
Send comments, questions, and suggestions to: macampbell@davidson.edu