This web page was produced as an assignment for an undergraduate course at Davidson College
Expression of My Favorite Yeast Genes: Fis1 & YIL067c
Figure 1: Color scale used to indicate expression in microarray experiments available at Expression Connection. Black indicates that expression under the experimental conditions was identical to expresion under control conditions. More vivid reds indicate greater levels of incuction relative to control conditions, and more vivid greens indicate greater levels of repression relative to control conditions (SGD, 2003; <http://genome-www4.Stanford.EDU/cgi-bin/SGD/expression/expressionConnection.pl>)
Expression Connection is a website that allows users to search for a particular gene of interest in many different microarray experiments. These experiments include the following:
* Evolution of expression during glucose limitation
* Expression during sporulation
* Expression during the cell cycle
* Expression during the diauxic shift
* Expression in response to DNA-damaging agents
* Expression in response to alpha-factor
* Expression in response to alpha-factor (various concentrations)
* Expression in response to environmental changes
* Expression in response to histone depletion
* Expression in response to varying zinc levels
* Expression regulated by the PHO pathway
Fis 1 Microarray Data
From web assignment 2, we learned that Fis1 is an s.cerevisiae protein involved in mitochondrial membrane fission. This process is imperative to mitochondrial function, which is to store energy from food as ATP and to distribute this energy to organelles.
When I searched Expression Connection for Fis1, the only available results were from the Evolution of expression during glucose limitation experiment. These results show that initially, Fis1 is repressed during glucose limitation, but that over time it is induced. As yeast realize that their glucose supply is being depleted, it would make sense for them to alter their methods of energy storage and distribution. Mitochondria need to change shape in order to change how they store and distribute energy, so Fis1 - a player in mitochondrial fission -is an important part of this process, explaining why it is induced during glucose limitation. The proteins that clustered with similar expression patterns are not grouped very convincingly; the gray boxes indicate that no data was available. Additionally, none of the proteins grouped with Fis1 are present on the outer mitochondrial membrane or have any known association with mitochondria. Also, out of the proteins listed, none are located in the same chromosomal region as Fis1, making it doubtful that any of these genes are regulated by the same promoter.
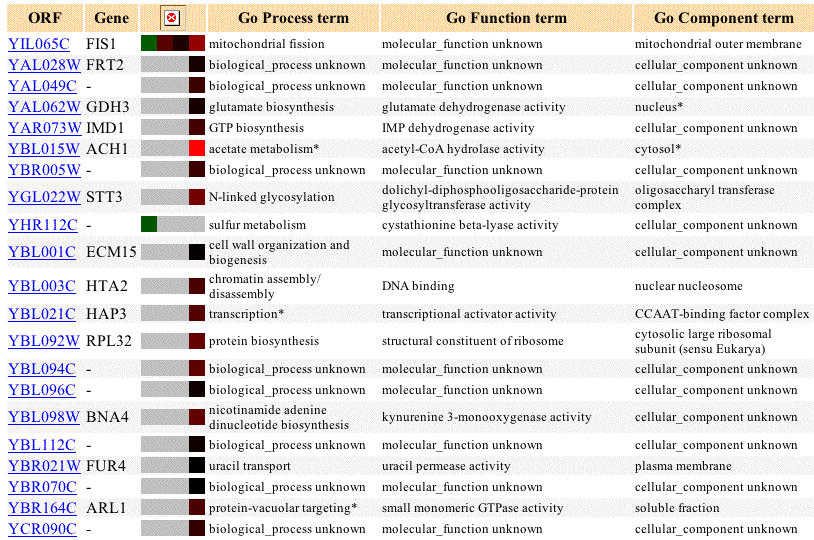
Figure 2: Evolution of expression for Fis1 and similarly expressed genes during glucose limitation. Data includes up to 20 similar genes, each with a Pearson correlation greater than 0.8 to Fis1 (SGD, 2003; <http://genome-www4.Stanford.EDU/cgi-bin/SGD/expression/expressionConnection.pl?orf=YIL065C&dataset=evolution&type=similar>).
Abstract for the glucose paper
Function Junction also provices a wealth of microarray data for genes of interest. When I searched for Fis1, the Yeast Microarray Global Viewer provided results from some 80-odd experiments. I was able to scale this amount down to experiments of greater interest by searching in the actual Yeast Microarray Global Viewer and setting the filter to 2. These results are pictured below. I have selected a few notable experiments to discuss in greater detail than shown in the figure below.
In the Causton Stress experiment, researchers observed expression levels in response to various environmental stresses. It can be seen that Fis1 was induced in response to many of these stresses. Why would remodeling of the motochondrial membrane, namely fission, be necessary to respond to stress? Perhaps the mitochondria must go through morphological changes in order to redistribute energy to new parts of the cell that require energy in times of stress.
In the Jelinsky MMS experiment, expression of Fis1 was induced in response to a DNA alkylating agent. It seems possible that the mitochondrial membrane would need to remodel in response to other genes being mutated.
The Kobor CTD experiment showed that Fis1 was widely repressed in experiments where genes involved in transcription with RNA polymerase II (which makes mRNA) were deleted or mutated. If mRNA is not made properly, protein synthesis is disruptively altered. Protein synthesis is vital to normal cell function, and is vital to the production of all proteins, including Fis1, which was most likely not produced in response to these experimental conditions.
In the Cohen YAPs, Fis1 was highly induced in response to mutations in Yap1, a transcription activator involved in response to environmental stresses. If a transcription activator in response to stress was deleted or mutated, Fis1 would be repressed, as it would probably be induced in response to stress, as we saw from the results discussed above from the Causton Stress experiment.
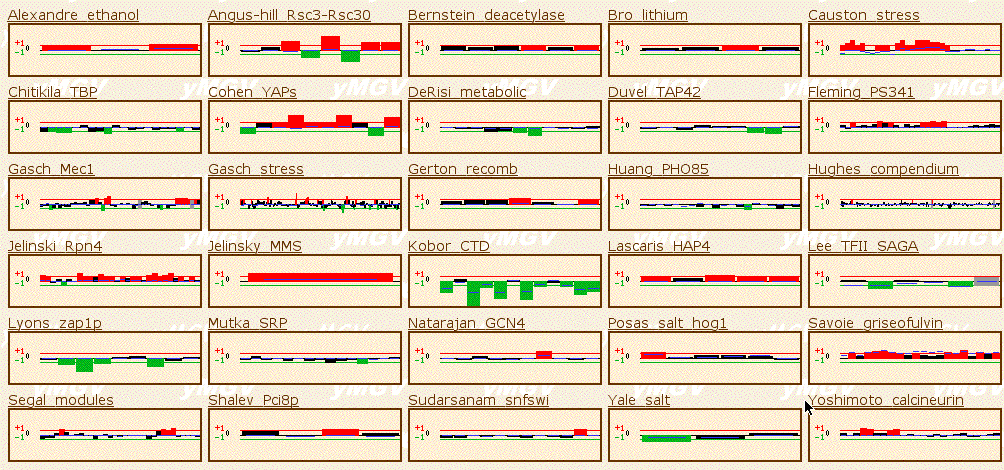
Figure 3: Yeast Microarray Global Viewer results for Fis1 with filter set to 2 (SGD, 2003; <http://transcriptome.ens.fr/ymgv/>).
Although not very extensive, microarray data for Fis1 gives support to its function as an outer mitochondrial membrane protein playing a role in energy storage and distribution.
YIL067C Microarray Data
Web assignment 2 attempted to make predictions about the function of YIL067c, a non-annotated yeast gene. My results were fairly vague, and my only conclusions were that it is an intermembrane protein with an LCCL domain that may be a region of functional importance in the way YIL067c folds. Hopefully microarray data will suggest a more specific functional role for YIL067c.
A search for YIL067C in the various experiments available at Expression Connection yielded (thankfully!) more results that my above search for Fis1.
The Evolution During Glucose Limitation experiment results for YIl067C are not conclusive. YIL067C's expression pattern during glucose deprivation shows no major differences from that of its expression under control conditions.
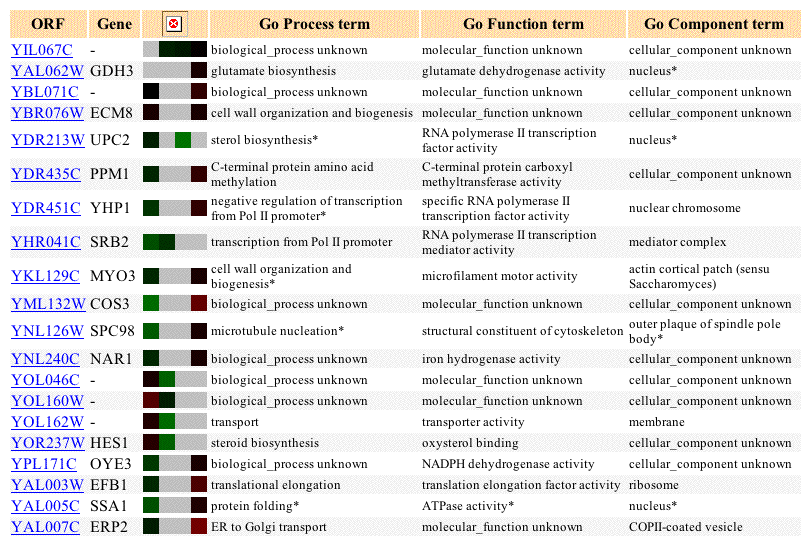
Figure 4: Evolution of expression for YIL067C and similarly expressed genes during glucose limitation. Data includes up to 20 similar genes, each with a Pearson correlation greater than 0.8 to YIL067C (SGD, 2003;<http://genome-www4.Stanford.EDU/cgi-bin/SGD/expression/expressionConnection.pl?orf=YIL067C&dataset=evolution&type=similar>).
Abstract for the glucose paper
When observing changes in gene expression during sporulation,
it can be seen that YIL067C is repressed at 2 hours but is induced later on,
most significantly at 7 hours. The known gene that YIL067C is expressed most
smilarly to during sporulation is YOR107W, which activates GTPases to play a
role in protein signaling. Many of these signaling events happen at the cell
membrane, suggesting that this protein might also be an intermembrane protein
like I have predicted for YIL067C. Based on expression patterns, YIL067C groups
with many proteins in the plasma membrane, as well as proteins in scattered
places like the mitochondrion, nucleus, and Golgi. Functionally, the proteins
it groups with are diverse and include proteins with transcription factor activity,
transporter activity, GTPase activity, ATPase activity, and protein kinase activity.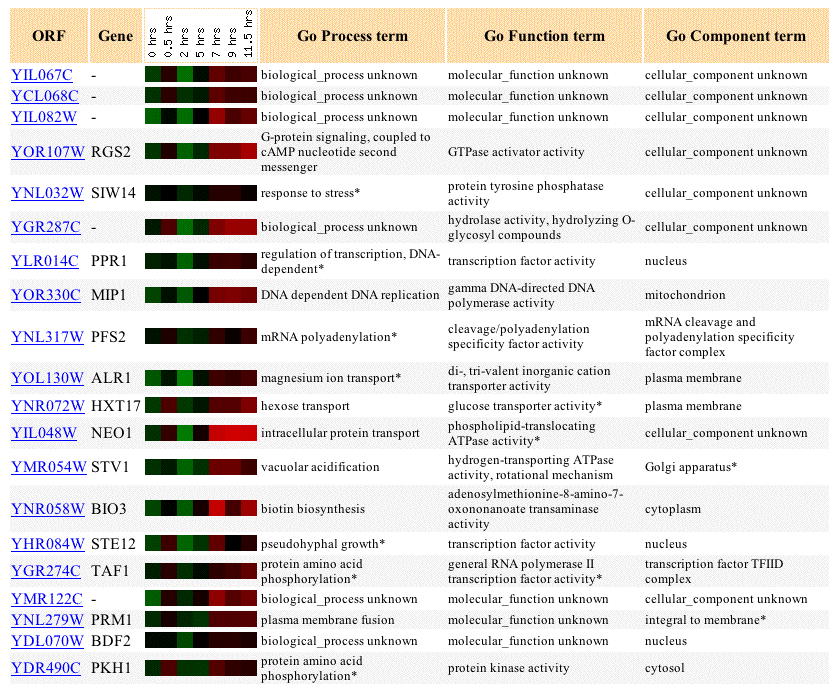
Figure 5: Expression of YIL067C as well as similarly expresed genes during sporulation. Data includes up to 20 similar genes, each with a Pearson correlation greater than 0.8 to YIL067C (SGD, 2003; <http://genome-www4.Stanford.EDU/cgi-bin/SGD/expression/expressionConnection.pl?orf=YIL067C&dataset=sporulation&type=similar>).
Abstract for sporulation paper
YIL067C is incuded and repressed to some degree at various points throughout the cell cycle. However, the low levels of induction and represson and the seemingly random times of induction and repression suggest that YIL067C plays little role in the cell cycle. It was also not expresed similarly to any other genes in this experiment.
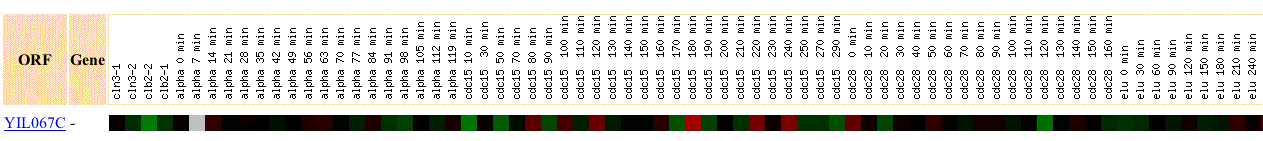 Figure 6: Expresion
of YIL067c during the cell cycle. Click on link in citation to view the four
columns of data unable to fit on the right side of this page (SGD, 2003; <http://genome-www4.Stanford.EDU/cgi-bin/SGD/expression/expressionConnection.pl?orf=YIL067C&dataset=cellCycle&type=similar>).
Figure 6: Expresion
of YIL067c during the cell cycle. Click on link in citation to view the four
columns of data unable to fit on the right side of this page (SGD, 2003; <http://genome-www4.Stanford.EDU/cgi-bin/SGD/expression/expressionConnection.pl?orf=YIL067C&dataset=cellCycle&type=similar>).
Abstract for Cell Cycle paper.
These results show that YIL067C is primarily repressed during the diauxic shift (when growth conditions are changed from anaerobic to aerobic), except for slight induction at 20.5 hours. This suggests that YIL067C is not involved in the conversion from anaerobic respiration to aerobic respiration. However, the genes that YIL067C clusters with may be helpful for my analysis. First of all, YIL067C clusters very closely with YIL082W, which it also clustered with in the sporulation experiment discussed above. This suggests that these two genes are regulated by the same mechanism and may share some similar function. Unfortunately, YIL082W remains a non-annotated gene so cannot help me determine a function for YIL067C. PPR1 is a known gene that clusters with YIL067C in both soprulation and diauxic shift experiments. The PPR1 gene product is located in the nucleus and is involved in transcription factor activity. Possibly YIL067C is part of the nuclear membrane and is involved in transcription in some way.
Other genes YIL067C clusters with in this microarray seem to be present in the nucleus, endoplasmic reticulum membrane, and cell membrane. The presence of membrane proteins in this cluster gives even more support to the prediction that YIL067C is an integral membrane protein. The function of many genes listed below is still unknown, but it seems like a good prediction that YIL067C is involved in transcription some level.
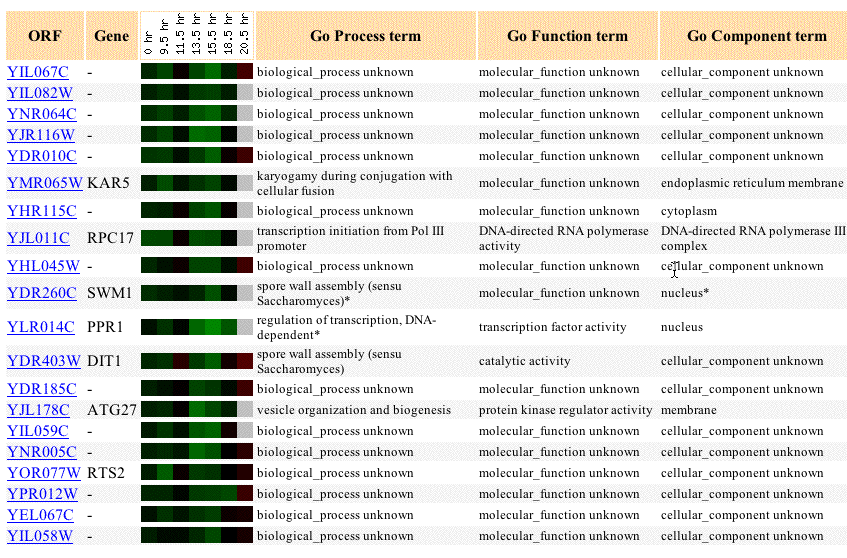
Figure 7: Expression of YIL067C as well as genes with similar expression during the diauxic shift. Data includes up to 20 similar genes, each with a Pearson correlation greater than 0.8 to YIL067C (SGD, 2003; <http://genome-www4.Stanford.EDU/cgi-bin/SGD/expression/expressionConnection.pl?orf=YIL067C&dataset=diauxic&type=similar>).
I also searched Function Junction's Yeast Microarray Global Viewer for YIL067C and found the results pictured below. I have selected a few notable experiments to discuss in greater detail than shown in the figure below.
In the Cohen YAPs, YIL067C was highly repressed in response to a certain mutation in Yap1, a transcription activator involved in response to environmental stresses. Repression in response to a transcription activator deletion would indicate that YIL067C did not need to be transcribed to help make up for Yap1's loss of function. Therefore, YIL067C probably does not play a similar role to Yap1. It may also be true that YIL067C's expresion is regulated by Yap1, which would explain its repression in response to Yap1's mutation.
The Kobor CTD experiment showed that YIL067C was strongly induced in one experiment where a gene (entitiled SRB10) involved in transcription with RNA polymerase II (which makes mRNA) was deleted or mutated. If mRNA is not made properly, protein synthesis is disruptively altered. Since YIL067C was incuded, this might suggest that it was transcribed to compensate for the lack of RNA polymerase II activity, suggesting a role in RNA synthesis. YIL067C may only be able to play a compensatory role for SRB10, as it was repressed in mutations in other similar genes in the Kobor CTD experiment.
In the Jelinsky MMS experiment, expression of YIL067C was induced in response to a DNA alkylating agent. Because I know very few specifics about YIL067C, this information is not very helpful.
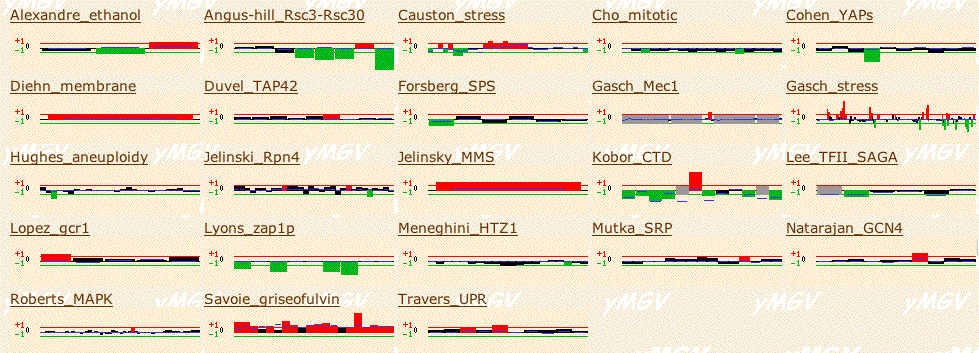
Figure 8: Yeast Microarray Global Viewer results for YIL067C with filter set to 2 (SGD, 2003; <http://transcriptome.ens.fr/ymgv/>).
Microarray data for YIL067C was not as helpful as I had hoped for predicting a function, but I still am confident that it is an intermembrane protein. Also, I have a few new predictions. By the guilt-by-association method, I feel like YIL067C may be integral to the nuclear membrane and may play a role in transcription. It might also be involved in signaling at either the cell membrane or the nuclear membrane.
References
Sacchromyces Genome Database. 2003. Expression Connection. <http://genome-www4.Stanford.EDU/cgi-bin/SGD/expression/expressionConnection.pl>
Sacchromyces Genome Database. 2003. Yeast Microarray Global Viewer. <http://transcriptome.ens.fr/ymgv/>
Questions? Comments?E-mail Sara