Expression Profiles of LAS17 and YOR186w
This web page was produced as an assignment for an undergraduate course at Davidson College.
In a previous assignment, I described my favorite Saccharomyces cerevisae gene and an unannotated proximal gene. I described Las17’s function and protein structure, and I hypothesized YOR186w’s function. In this assignment, I will use DNA microarrays to further describe these genes’s expression pattern and further test my hypothesis of the function of YOR186w. |
||
DNA Microarrays |
||
DNA microarrays are a useful new technology that allows researchers to examine the expression patterns of genes within a cell under different conditions. The expression pattern describes the amount of transcription of a certain gene within a cell. This technique allows us to study the genome as it responds to different stimuli, and thus, discover or better understand genes within the context of an environmental response by the cell. If you are comparing two genes with microarrays, one genome would be dyed with red and the other would be dyed with green. If both genomes were transcribing the same genes, the spot would turn yellow indicating both red and green were present within that microarray sample. Furthermore, computer analysis programs exist to allow researchers to determine the ratios of which genes are transcribed in different genomes. Researchers are also using DNA microarrays to infer gene function of unannotated genes, confirm gene function of annotated genes, and understand pathways and genes involved in cellular processes. Data from microarrays are described along a color gradient. Typically, green represents repression and red represents induction of a gene. The intensity and color of the resultant spot is due to the ratio of change from a standard condition to the experimental condition such as heat shock. From these data, researchers are also attempting to cluster genes. Clustering is a process by which genes with similar expression patterns are graphed and assigned a correlation coefficient. Based on these correlation coefficients, researchers use clustering methods to determine which genes are more closely related in expression patterns. From these analyses, researchers can develop dendrograms that provide similar information as a phylogenetic tree. |
||
Fig. 1. This is the color scale for these microarrays where Cy3-labeled DNA is expressed with a green color. Conversely, Cy5-labled DNA is expressed with a red color. Black means that the gene either had no change in expression or the gene was not transcribed in high enough levels to be detected. |
||
Las17/BEE1 |
||
Las17 is involved with actin assembly within S. cerevisae located on chromosome 15. More specifically, this gene regulates Arp 2/3, which is responsible for allowing actin polymerization by facilitating the initial attachment of a new actin polymer to the original strain. Furthermore, Las17 is a known homolog for a human disease known as Wiskott-Aldridge Syndrome that is characterized by a loss of actin function in leukocytes of primarily men. Mutations of this gene are known to affect endocytosis and cellular budding. |
||
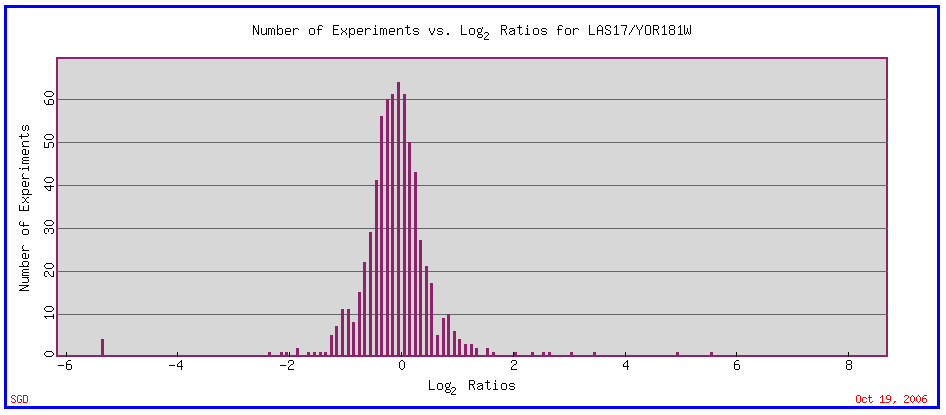
Fig.1. The general expression ratios of Las17. "LAS17 Expression Connection". SGD. 2006. http://db.yeastgenome.org/cgi-bin/expression/expressionConnection.pl |
||
Similarly expressed genes: Expression in response to environmental changes
This study examined the effects of stressful environmental changes such as heat shock on the mRNA expression levels within yeast cells. Overall, a large gene set (approximately 900 genes) were implicated in reactions to most stressful conditions. In the figure below, we can see the expression levels of Las17 during stressful conditions. |
||
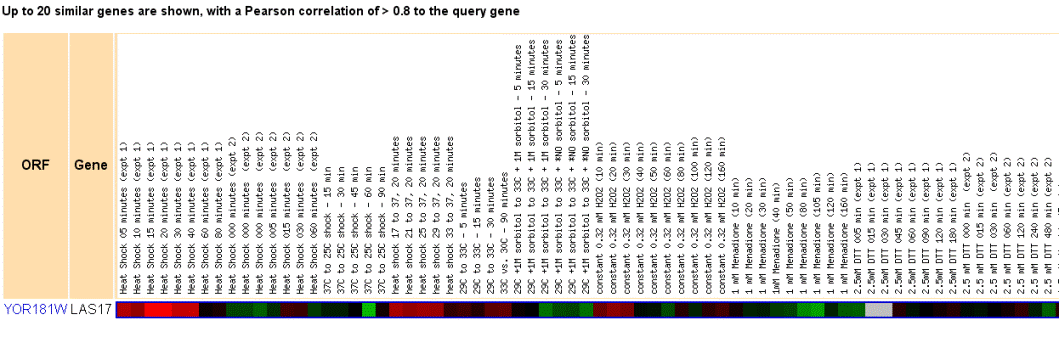
Figure 3. The expression profile of Las17 during a series of stressful environmental conditions. Las17 appears to be greatly induced during periods of heat shock as can be seen in the first set of experiments and again at different temperature ranges. One possible explanation for this reaction given Las17's known function is that at higher temperatures yeast cells grow more quickly or replicate more quickly. Thus, at higher temperatures, genes required for actin polymerization yielding growth and facilitating budding would be activated as well. However, since the results of this experiment appear to be somewhat mixed (see the somewhat repressed region in the second heat shock experiment), I would assume that Las17 is involved in growth reactions or endocytosis reactions to heat shock, especially at lower temperatures. Furthermore, in the cold shock experiment, changing the temperature from 37C to 25C, Las17 appears to be expressed at low levels and is strongly repressed in one reaction. |
||
Similarly Expressed Genes: Expression in Response to Histone Depletion
Histones are proteins within eukaryotic nuclei that assists with chromosome organization and perhaps gene regulation. This study shows that histone deletion from the nucleus produces a change in gene expession in 25% of yeast genes. In the figure below, we see the twenty genes that have similar expression profiles to Las17 when one part of the histone complex is deleted from the cell. |
||
Fig. 4. The expression profile of 20 genes in response to histone deletion. The figure above shows that initially following histone deletion, Las17 is strongly repressed before being strongly induced after the initial reaction to histone deletion. Other genes with similar expression patterns are also responsible for factors involved with endocytosis throughout the cell such as YAP1801. Furthermore, expression patterns of Las17 are very closely matched with expression patterns of genes such as MRE11 responsible for telomere maintenence. Thus, Las17 may be involved with assisting chromosome organization by creating more actin chains to support chromosomes.
Fig. 5. We can see a general increase of LAS17 induction as more histone is depleted. |
||
Similarly Expressed Genes: Expression During Sporulation
This experiment examined gene expression of yeast during sporulation. Sporulation consists of meiosis and then spore morphogenesis. Spores are formed when diploid cells are introduced to stressful conditions. Sporulation will not occur in yeast cells that are haploid or polyploidy. The representation of expression profiles below shows Las17 and other genes with similar expression patterns. |
||
Fig. 6. The expression profile of 20 genes in response to the yeast process of sporulation. Las17 appears to be strongly represed as the process of sporulation progresses. Initially, I was a bit surprised by these results because I would assume that actin would be required during sporulation to ensure that all of the proper elements are in the correct location for successful sporulation. However, repression appears to occur towards the end of the sporulation process when perhaps the yeast cell wants to separate the spores, and hence, the cell represses Las17 and other actin polymerization genes to ensure that the spores separate and are not reattached by increased actin production. Furthermore, the cell may be simply shutting down various cellular processes towards the end of sporulation because as we can see above, several genes from a variety of functions are also repressed only at the end of the sporulation process.
Fig. 7. We can see an increase of gene repression throughout the process of sporulation. |
||
Similarly Expressed Genes: Expression During the Unfolded Protein Response This study examined the yeast cellular mRNA expression levels during the unfolded protein response that occurs within the endoplasmic reticulum. As proteins are produced, they travel through the ER to complete protein development such as tertiary or quaternary structures. However, if the ER is stressed, some proteins may not develop their tertiary or quaternary structures properly. Thus, the cell has developed two processes for eliminating these non-functional proteins. One method is exocytosis from the ER into the cytoplasm where the protein is degraded. Another method is the unfolded protein response in which a pathway is utilized to slowly degrade the protein within the ER. Below is the expression profiles of genes that may be involved with this process. |
||
Fig. 8. Expression profile of 20 genes induced or repressed during different methods of protein degradation. In the figure above, as expected, during exocytosis into the cytoplasm for protein degradation, Las17 is strongly induced. This is an expected result because actin is known to function in endocytosis and exocytosis since these processes require movement and production of new actin polymers. Furthermore, we see a repression of Las17 during the unfolded protein response. While I did not expect Las17 to have a strong induction ratio, I was surprised to see that the gene appears to be actively repressed during this process. One explanation of these results is that actin production may disrupt the unfolded protein response within the ER. |
||
YOR186w |
||
YOR186w is an open reading frame on chromosome 15 that has no known biological function, molecular function, or cellular component. In my previous assignment (LINK), the best sequence similarity I observed by database searching was for a gene named FLiP that is located on a plasmid within S. cerevisae cells. FLiP promotes duplication events within the plasmid, and thus, I suggested that perhaps this open reading frame may have be incorporated into the S. cerevisae genome and may or may not be functional within this genome. In this assignment, I will use microarrays to prove or disprove my hypothesis and if possible form a better or more detailed hypothesis via “guilt by association”. We know little about this gene, but if its expression pattern is similar to a group of genes, we may be able to infer that its function is similar to the genes of known function that YOR186w clusters with. |
||
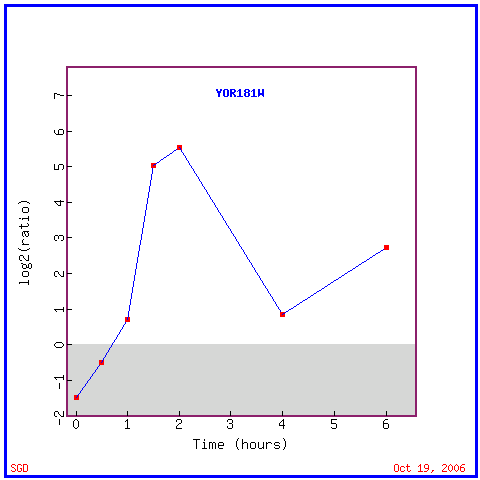
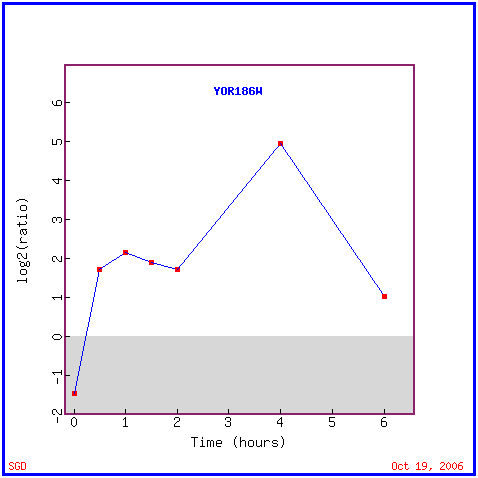
Fig. 9. Graphical representation of expression changes in experiments studying YOR186w. "YOR186w Expression Connection". SGD. 2006. http://db.yeastgenome.org/cgi-bin/expression/expressionConnection.pl |
||
Similarly Expressed Genes: Expression During the Unfolded Protein Response This study examined the yeast cellular mRNA expression levels during the unfolded protein response that occurs within the endoplasmic reticulum. As proteins are produced, they travel through the ER to complete protein development such as tertiary or quaternary structures. However, if the ER is stressed, some proteins may not develop their tertiary or quaternary structures properly. Thus, the cell has developed two processes for eliminating these non-functional proteins. One method is exocytosis from the ER into the cytoplasm where the protein is degraded. Another method is the unfolded protein response in which a pathway is utilized to slowly degrade the protein within the ER. Below is the expression profiles of genes that may be involved with this process. |
||
Fig. 10. The expression profiles of genes similar to YOR186w during the unfolded protein response. Similar to Las17, YOR186w appears to be involved in the process of degrading unfolded proteins in the cytoplasm. Several of the genes that have similar expression profiles appear to interact with mRNA and DNA such as MSS11 or NUP2. These genes appear to regulate transcription and assist mRNA export from the nucleus. |
||
Similarly Expressed Genes: Expression in Response to Peroxisome Induction and Repression
This experiment examined the gene expression changes as researchers manipulated the quantity of peroxisomes within the cell. First they induced the level of peroxisomes over time, tested one trial of reducing the number of peroxisomes, and final tested a control. The oleate increases the amount of peroxisomes within the cell, glycerol reduces the number of peroxisomes, and glucose does not alter the amount of peroxisomes within the cell. Below are the results of this study. If genes were induced during the first trial, they should then become repressed during the second trial. |
||
Fig. 11. The expression profile of genes similar to YOR186w with a Peason correlation coefficient of at least 0.8. As expected genes that were induced in conditions with many peroxisomes were then repressed when they experienced few peroxisomes indicating that gene expression of these genes is linked in some way with peroxisome production and function. Once again, YOR186w appears to cluster with genes that are involved with DNA and RNA. |
||
Similarly Expressed Genes: Expression in Response to Histone Depletion
Histones are proteins within eukaryotic nuclei that assists with chromosome organization and perhaps gene regulation. This study shows that histone deletion from the nucleus produces a change in gene expession in 25% of yeast genes. Below we see the reactions of YOR186w and other genes with similar expression patterns to histone depletion. |
||
Fig. 12. The expression profiles of genes similar to YOR186w in response to histone depletion. This grouping of expression profiles is extremely striking. YOR186w appears to have nearly the exact expression profile as BTN2, involved with amino acid transport. Furthermore, YOR186w also has a similar expression profile as BNA4, involved in NAD biosynthesis.
Fig. 13. We see an induced expression of YOR186w over time until after 4 hours when the level of induction begins to decrease. |
||
Predicted Function of YOR186w |
||
| Due to the expression profiles above and the theory of guilt by association, I refute my earlier hypothesis that YOR186w is involved with duplication within the yeast genome. Instead, I now feel that this unannotated gene may instead play a role in gene transcription or translation. In figure 10, we determined that YOR186w has expression profile similarity with the gene responsible for regulation of the RNA polymerase promoter and the gene involved with protein biosynthesis. In figure 11, YOR186w shared similar expression profiles with genes responsible for ribosome biosynthesis, amino acid phosphorylation, and transcription. Finally, in figure 12, YOR186w shared similarity with amino acid transport and NAD biosynthesis. Due to guilt by association, I conclude that YOR186w may function in cellular translation and may be involved with the movement and regulation of amino acids as they form proteins. | ||
| Citations | ||
|
"LAS17 Expression Connection". SGD. 2006. http://db.yeastgenome.org/cgi-bin/expression/expressionConnection.pl "YOR186w Expression Connection". SGD. 2006. http://db.yeastgenome.org/cgi-bin/expression/expressionConnection.pl |




Questions? Please contact Krcecala@davidson.edu
© Copyright 2006 Department of Biology, Davidson College, Davidson, NC 28035