
This web page was created for an undergraduate assignment at Davidson College.
Stella Kenyi
BIO 307: Immunology
Dr. A. Malcolm Campbell
CD45 is a protein tyrosine phosphatase (PTP) located in hematopoietic cells except ethrocytes and platelets. CD45 is also called the common leukocyte antigen, T220 and B220 in mice. The protein tyrosine kinases constitute a family of receptor-like and cytoplasmic inducing enzymes that catalyze the dephosphorylation of phosphostyrosine residues and are characterized by homologous catalytic domains ( Barford et al. 1994). CD45 is a protein that has several isoforms and the hematopoietic cells express one or more of the isoforms. The specified expression of the CD45 isoforms can be seen in the various stages of differentiation of normal hematopoietic cells ( Virts et al. 1997). CD45 is uniformly distributed in plasma membrane and enrich regions of T cell and B cell contact. In fact, CD45 constitute 10% of the molecules on the surface of expressing cells( Katz 2003 ). Some isoforms of CD45 are CD45RO, CD45RA, and CD45RB. Each CD45 isoforms is distinguished from one another isoform depending on the type of exon the CD45 has or the exons the CD45 does not have. The CD45RA isoform contains the A exon only and the CD45RB has the B exon only whereas the CD45RO has neither the none of the exons: A, B, or C.
CD45 is made of a long single chain type one transmembrane protein with approximately 1100-1300 amino acids. The extracellular domain of this protein contains amino acids that range from 300-500 amino acids and a long cytoplasmic tail with approximately 700 amino acids that contains repeats for the two phosphatase domains. Most researchers note that the first domain has more activity compared to the second domain and this is generally believed to be true since the second domain does not have necessary amino acids pertinent for catalytic activity. The amino acids in the second domain are changed in comparison to the first domain (Okumura et al. 1996). Three fibronectin type III domains are believed to make up the extracellular domain ( Okumura et al. 1996). As noted above, Each CD45 contains one of the two exons exons: A, B, and C and the CD45 termed the CD45RO contains neither of the exons. The exons determining what type of CD45 is found on the plasma membrane are located in the N terminus of the CD45 protein. Similarly as noted above, CD45 is a protein tyrosine phostphatase that functions to regulate Src kinases required for T and B cell receptor signal transduction ( Kung et al. 2000). The CD45 dephosphorylates a negative regulatory residues on one or more of the protein tyrosine kinases involved in receptor mediated second messenger formation (Donovan and Korezky 1993). In T cells, the Src family kinases such Lck, and Fyn, are molecules regulated by CD45 and in B cells the Src family kinases such as Lyn and Blk. In T cell and B cells, the phosphorylation of the ITAMS is an intracellular signal that alerts the cell that the lymphocyte has detected it specific antigen. The ITAMs are immunocreceptor tyrosine bases motifs (ITAMS) composed of two tyrosine residues separated by amino acids. When ITAMS’s tyrosines are phosphorylated by the receptor associated tyrosine kinases, the ITAMS are now able to bind second family protein tyrosine kinases such as CD45 . The second family protein kinases’ SH2 domains now have a high affinity for the ITAMs and this leads to further phosphorylation( Janeway et al. 2001). In T cells, CD45 in particular phosphorylates Csk, an inhibitory protein tyrosine kinase therefore the activation is particularly important because it controls levels of tyrosine activity in lymphocytes. In B cells , Ca2+ is transduced by the B cell receptor and this signaling is believed to induce the expression of CD45 ( Justment et al. 1991). Scientist also use CD45 to detect naïve T cells from effector cells and memory T cell. Different proteins are expressed during differentiation of T cells and B cells. Using flow cytometry, Scientist can measure relative expression levels of CD45 isoforms , a marker of naïve versus memory CD4+ T cell status ( Tan et al. 2002).
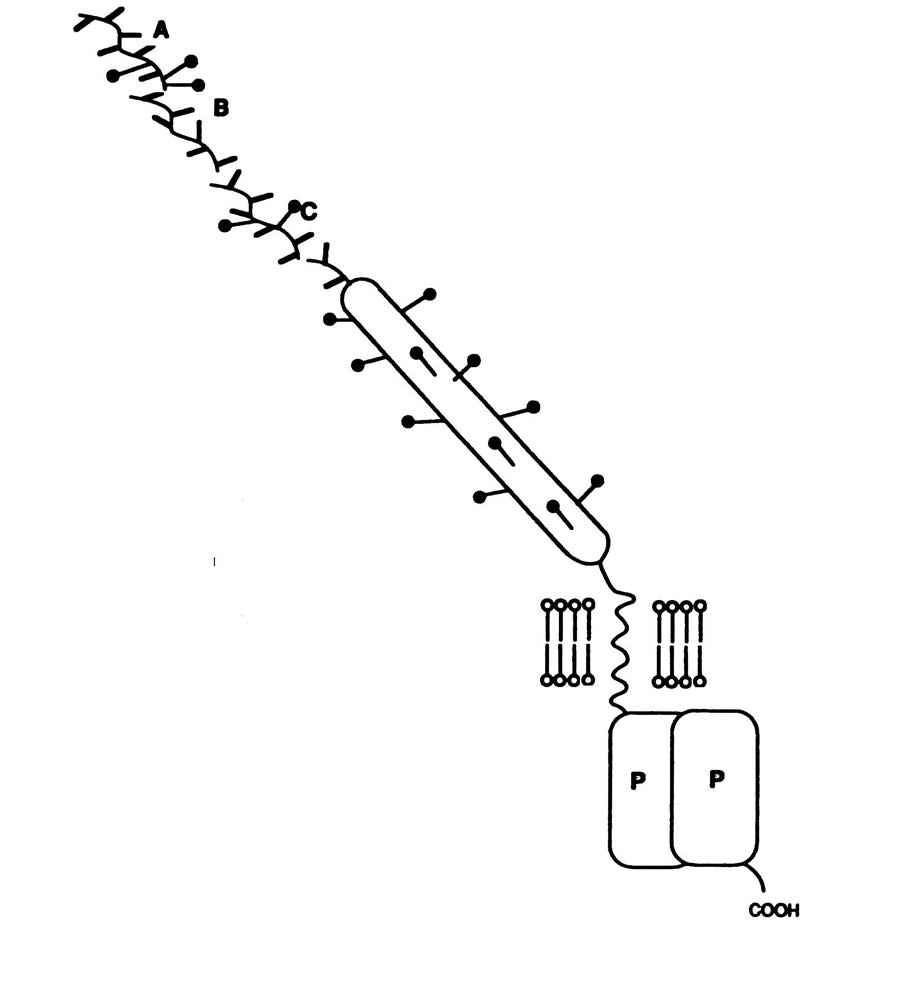
Figure 1. CD45 in plasma membrane. The figure shows a CD45 molecule, which is a tyrosine protein kinase, embedded in the plasma membrane. The exons A, B, and C are located in the extracellular domain and the protein kinase domain is located in the intracellular side. ( Permission requested from A. Neil Barclay found on Ph.D 03/20/03). A. Neil Barclay
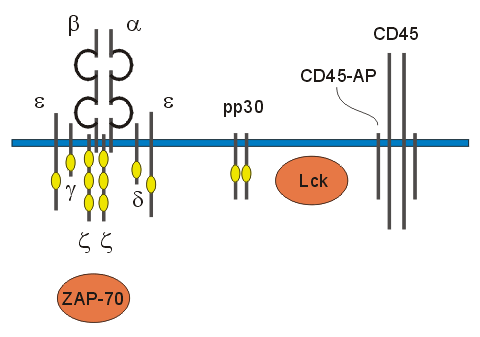
Figure 2. Signal transduction through the T cell receptor. T cell receptor complex with the a and b and the CD3. The CD3 is made of g, d, and two e chains. The T cell receptor complex has ITAMs and the phosphorylation of the tyrosine residues on the ITAMS alerts the T cell of antigen binding. The phosphorylation leads to further phosphorylation of second family tyrosine proteins such as CD45, ZAP 70 and Lck. The phopshotidlyinostisol 3 can also be recruited ( Permission received from Dr. Kevin Mackenzie Ph.D on 3/20/03). Interactive model from website
As noted earlier,
humans have CD45 missing from their erythrocytes and their platelets
but they are readily available on the rest of the hematopoeitics cells. A
mutation in CD45 may lead to dysfunction on B lymphocytes and low T cells;
therefore causing immunodeficiency. Before scientists had no reports to
knowledge of a human deficiency but recent experiments such as with
a male patient having one deletion at one allele and a point mutation at
another allele( Kung et al. 2000) proved otherwise. SCID appeared in the patient after a two
month period and the T lymphocytes are
diminished and unresponsive to mitogen stimulation and the B cell
decreased as the patient aged. This suggested to the scientists that CD45
deficiency is linked and results in T lymphocytes and B lymphocytes dysfunction
( Kung et al. 2000). Therefore missing CD45 may result in nonfunctioning T
lymphocytes and B lymphocytes. In CD45 mutant cell lines, CD45-deficient mice
and CD45-deficient human SCID patients, CD45 is
required for signal transduction through antigen receptors (Sasaki et al.1995). Research
has also investigated and showed that a point mutation in the CD45 in the heterozygous
state of multiple sclerosis. A nucleotide transition in exon 4 of the gene locus
interferes with mRNA splicing and results in altered expression of CD45
isoforms on lymphocytes (Jacobsen 2000). The Scientists found a close
association with disease and three multiple sclerosis nuclear families. As
noted above, Scientist used a marker of naive versus memory T cells. Patients
with various clinical diagnosis showed such as Alzheimer's disease and dementia
showed lower levels of CD45RA and an increase in the CD45RO( Tan et al.
2002). The lymphocytes lost the CD45 exons such as A, B, and C; therefore most
CD45 proteins became CD45RO. The lymphocytes of newborns with different
variation in infection and infants with no infection show different ranges of
CD45RO; the Infants with definite infection had 10% more CD45RO than infants
with no infection (Tezuka et al. 1998).
When researches perform allografts, target CD45RB by creating an anit-CD45RB antibodies which allow the tolerance for the allografts. The researchers show that anti-CD45RB treatment alters CD45 isoform expression on T cells, which is associated with rapid up-regulation of Cytolytic T lymphocyte–associated antigen 4 (CTLA-4) expression. These effects appear specific and occur without up-regulation of other activation markers ( Fecteau et al. 2001). Therefore alteration of the isoform allow for change to a different isoform and this change allows for the allograft tolerance.
REFERENCES
Braford, D., Flint, A.J., Tonks, N.K., Crystal structure of human phosphotase 1B. 1994. Science 11:253(5152):1373.
Donavan, J., and G.A., Koretzky.1993. CD45 and the immune response. Journal of American Society of Nephrology. 4(4) 976-985.
Fecteau, S., Badasodonna G.P., Freitas, A., Ariyan, C., Sayegh.M.H., and D.M. Rothestein.2001.CLTA-4 up-regulation plays a role in tolerance mediated by CD45. Nature- Immunology 2: 58-63.
Kung, C., Pingel, J.T., Heikinheimo, M., Klemola, T., Varkila, K., Yoo, L.I., Vuopala, k., Poyhonen, M., Rogers, M., Speck, S.H., Chatilla, T.A., and T.M. Matthews.2000. Mutations in the tyrosine phosphotase CD45 genes in child with sever combine immunodeficiency disease.Nature-Medicine.
Jacobsen, M., Schweer, D., Zeigler, A., Garber, R., Schock, S., Schwinzer, R., Wonigeit, K., Lindert, R.B., Kantraci, O., Schaefer-Kline, J., Schipper, H.I., Oretel, W., Heindenreich, F., Weinshenker, B.G., Sommer, N., and B. Hemmer. 2000. A point mutation in PTPRC is associated with the development of multiple sclerosis. Nature-Genetics. 23(4) 495-499.
Janeway, A.C, P. Travers, M. Walport., J.M. Shlomchik.2001. Immunology: The Immune System in Health and Disease. New York, NY: Elsevier Science Ltd. Garland Publishing, pp 196, 416-417.
Justement, L.B., Campbell, K.S., Chein, N.C., and J.C., Cambier. 1991. Regulation of B cell antigen receptor signal transduction and phosphorylation by CD45. Science. 252(5014):1839-1842.
Katz, K.S., Protein Preview on the Web CD45.< http://www.ncbi.nlm.nih.gov/prow/cd45/cd45.html > Assessed 3/16/03.
Okumara, M., Matthews R.J., Robb, B., Litman G.W., Bork, P., and M.L. Thomas. 1996. CD45 extracellular domain sequences from divergent vertebrate suggests the conservation of three fibronectin type III domains. Journal of Immunology.157:1569.
Sasaki Y, Hakusui H, Mizuno S, Morita M, Miya T, Eguchi K, Shinkai T, Tamura T, Ohe Y, Saijo N. 1995. A pharmacokinetic and pharmacodynamic analysis of CPT-11 and its active metabolite SN-38. Journal of Cancer Research 86:101-110.
Tan, J.A., Town, T., Abdullah, L., Wu, Y., Placzek, A., Small, B., Kroeger, J., Crawford, F., Richards, D., and M. Mullan. 2002. CD45 isoform alteration in CD+ Tcells as potential diagnostic marker of Alzheimer's disease. 2002. Journal of Neuroimunology. 132(1-2)164-172.
Tezuka, T., Sugita, K., Mizobe, N., Miyamoto, N., Nakamura, M., Kagami, K., Yokoyama, T., and S, Nakazawa. 1998. Transient increase in CD45RO expression on T lymphocytes in infected newborns. Pediatric Research 43(2) 283-290.
Virts, E., Barritt, D., Siden, E., and R.A., William. 1997." Murine mast cell s monocytes express distinctive sets of CD45 isoforms. Immunology 34(16-17) 1119-1197.