This page was created as an assignment for an undergraduate class at Davidson College. It is not intended to be used as medical advice.
THE IMMUNOLOGY OF LEPROSY
Leprosy, also known as Hansen’s disease, continues to be a major public health concern in many countries. There are today effective treatment strategies and modern medicine allows the disease to be effectively controlled, but public health barriers such as misdiagnosis and poorly trained professionals as well as social barriers such as the unwarranted but widespread stigma surrounding the disease have posed significant challenges to its control. Elimination of the myth of leprosy as an incurable, highly contagious, and debilitating disease has to change in order to encourage patients to seek treatment and diminish their discrimination from the community.
In 1991, the World Health Organization adopted an initiative to eradicate Hansen’s disease worldwide by the year 2000, with eradication defined as a prevalence of less than one in ten thousand persons. The number of Hansen’s disease patients dropped from 5.2 in 1985 to just 286,000 at the end of 2004. There has been a twenty percent annual decrease in the number of cases detected globally for the past five years. While the campaign was a success on a global level, eleven countries failed to meet this goal by the intended date and nine countries have still not eradicated the disease. These nine countries account for over seventy-five percent of leprosy cases. Brazil, India, Mozambique, Nepal, and Madagascar are currently the principal focus of elimination work (WHO 2005). Efforts are now being directed at implementing effective public health programs aimed at getting information to the poor people of endemic areas that are most likely to contract the disease and ensuring free treatment availability.
Leprosy is caused by infection with Mycobacterium leprae. These bacteria infect macrophages and Schwann cells, leading to skin lesions and peripheral nerve degeneration. If left untreated, permanent skin and nerve damage can result along with deterioration in the limbs and eyes. Contrary to popular thought, leprosy is not highly communicable. Prolonged contact with patients who have open lesions and are not in treatment is generally required for infection. The incubation period of the bacterium is generally about five to seven years, although symptoms can take as long as twenty years after the initial infection to appear (WHO 2005).
Leprosy manifests itself in two different clinical forms. Tuberculoid leprosy is the more mild form of the disease, in which mycobacterial proliferation and destruction is relatively contained by a cell-mediated immune response. This form of the disease is also referred to as paucibacilar Hansen’s disease, because patients exhibit on average one to five skin macules. Tuberculoid leprosy is rarely fatal. Lepromatous leprosy is a much more severe manifestation of the disease. Humoral immune responses predominate in the lepromatous form and are ineffective at containing the infection. M. leprae replicate freely inside macrophages and provoke extensive tissue damage. Lepromatous leprosy is also referred to as multibacilar Hansen’s disease and is characterized by numerous skin lesions, nodules, and plaques, a thickened dermis, and congestion of the nasal mucosa (CDC 2005).
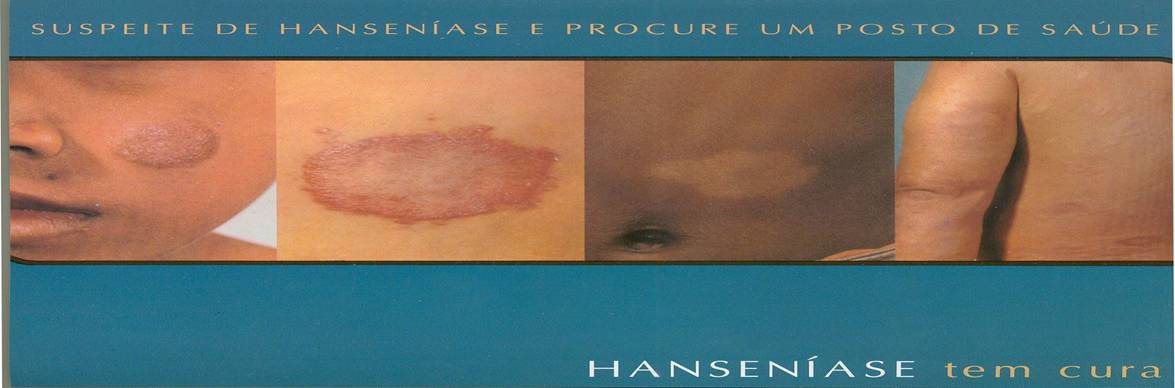
The key indicators of the early stages of leprosy are the spots that appear on the skin of patients. These spots may appear white, red, or copper-toned, may be raised or flat, and have no characteristic shape associated with them (see Fig.1). This variety poses a significant challenge to the diagnosis of the disease, particularly in regions where the public health workers may not be well-trained. The factor that makes leprosy distinguishable from the plethora of other diseases it could be confused for is the fact that skin spots do not hurt or itch, but are insensitive to tactile or thermal sensation due to peripheral nerve damage. Diagnosis is therefore made by conducting sensitivity tests, where the patient is asked whether they can feel certain stimuli. A more ideal diagnostic method would be to conduct a skin smear. These ensure that cases of multibacilar Hansen’s disease are distinguished from paucibacilar Hansen’s disease and the correct treatment strategy is implemented. However, sufficient laboratory resources are not available in most highly endemic regions.
The gross deformities frequently associated with ‘lepers’ are in fact only a risk if the disease goes undiagnosed and untreated for prolonged periods of time. Death of peripheral nerves can make it impossible to use certain appendages, and muscle atrophy follows. Bone and cartilage, although not the main centers of infection, can eventually deteriorate in response to M. leprae, leasing to misshaping of limbs. Such complications are not common with patients undergoing treatment and many therapeutic remedies are available to prevent them.
There is a very high incidence of ocular involvement among patients who suffered from Hansen’s disease resulting from infiltration of the tissues and damage to the nerves. Complications appear to be directly correlated with the duration of the disease. Most patients who had been infected with the disease for over fifteen years showed some sort of ocular involvement. Presentation ranged from cataracts, conjunctivitis, and complications of the lacrimal system to blindness (Wani et al., 2005).

Fig. 1. This pamphlet is an example of those used in government information campaigns in endemic regions of Brazil. It shows photos of several different possible dermatological presentations of the disease, outlines the symptoms, and directs people to go public health posts to receive free treatment. Perhaps more importantly, it emphasizes that patients with Hansen’s disease do not need to interrupt their normal activities to seek treatment (Prefeitura da Cidade Rio: Secretaria de Saude, personal property).
M. leprae was the first bacteria discovered to cause disease in humans. It was isolated in 1873 by the Norwegian physician Gerhard Henrik Armauer Hansen (WHO, 2003). Leprosy was renamed Hansen’s disease in his honor to counteract the growing stigma associated with the term ‘leprosy.’
M. leprae is an acid-fast, rod-shaped bacterium with a gram-positive cell wall. It is an obligate intracellular bacterium; therefore it can never be grown in a cell culture or agar solution (CDC). This bacterium preferentially infects macrophages and Schwann cells, leading to the pathology described above.
When a patient with active Hansen’s disease talks, coughs, or sneezes, they release droplets that contain the bacilli into the air. Most of these droplets don’t lead to active infection, even when inhaled by another person. Many are relatively large in size (>5μm in diameter), which can prevent them from reaching the small air sacs in the lungs, and instead they settle in the higher regions of the respiratory tract. Many of the body’s defenses can act in this region to eliminate bacilli. On the other hand, the moisture surrounding some droplets may evaporate quickly in the air, leaving light residues called droplet nuclei that are small enough to reach the alveoli (1-3μm in diameter) (Nardell, Piessens 2000). From the respiratory tract, M. leprae can invade the bloodstream and travel to areas of characteristic infection.
Several mechanisms have been proposed by which mycobacteria invade host macrophages. Opsonization of mycobacteria with C3 is possible through the alternative pathway of complement activation. The coating is then recognized by the complement receptors CR1, CR3, and CR4 and the mycobacteria are engulfed into phagosomes. Pathogenic bacteria appear to be unique in their use of the C2a complement fragment. Mycobacteria-associated C2a was shown to form a C3 convertase, resulting in mycobacterial opsonization and recognition by macrophages (Schorey et al., 1997).
Mycobacteria are engulfed by macrophages but are able to survive and become pathogenic by preventing the fusion of phagosomes and lysosomes. The mechanism for this includes blocking Ca 2+ signals and blocking the synthesis of proteins responsible for this fusion (Robbins 2005). When the macrophage is recognized and activated by a T-helper1 cell, the lysosome fuses with the phagosome and the mycobacteria are exposed to proteases that produce peptide fragments that can be bound to MHC class II molecules and presented at the cell surface to CD4 T cells.
Granulomas frequently result in patients with mycobacterial infections. These are sites of local inflammation characterized by a central aggregation of infected macrophages surrounded by T cells. Granulomas are useful to isolate pathogens and subject them to some degree of regulatory control from the surrounding T cells (Janeway et al., 2005).
Recombinant antigens, or stress proteins, are often produced by pathogens in response to the host environment. These are involved in eliciting an immune response and serve as important immune targets. Patients with tuberculoid leprosy show increased levels of antibodies against the 18 kDa and 28 kDa antigens, while patients with lepromatous leprosy exhibit antibodies against the 65 kDa antigen (Mohanty, 2004). It is speculated that mycobacterial presentation of different stress proteins could play a role in directing the type of immune response developed.
The paradigm that antigen presentation involves peptide presentation on MHC molecules is not all-inclusive. Lipid antigens can be presented by CD1 molecules in an MHC-independent antigen presentation pathway. CD1 molecules are roughly homologous to MHC molecules, but they are encoded on chromosome 1 and not as part of the major histocompatibility complex on chromosome 6; they are also nonpolymorphic. CD1b molecules present lipid components of mycobacterial cell walls to double-negative (CD4 -8 -) T cells. This mechanism allows for T cell recognition and killing of infected cells. CD1 proteins are highly expressed in patients with tuberculoid leprosy (Sieling et al., 1995).
Structures of lipid antigens do not vary as much between microbial species because they are essential to the integrity of the bacteria’s cellular envelope and could therefore be important antigens in the development of a specific immune response against mycobacteria. T cells from lepromatous patients exhibit much more limited responsiveness to lipid antigens than tuberculoid patients because of their limited expression of CD1 molecules. It is suspected that elimination of cross-reactive CD1-restricted T cells occurs in the periphery of patients with lepromatous leprosy, resulting in an unresponsive and ineffective subset of T cells in these patients (Sieling et al., 2005).
When an undifferentiated CD4 helper T cell recognizes the antigen presented on the MHC class II molecules of infected macrophages, the cytokine environment determines whether it will differentiate into a T-helper1 or T-helper2 cell. IL-12 and IFN-γ promote differentiation of T-helper1 subtypes, leading to a cell-mediated immune response and the less severe tuberculoid manifestation of the disease. T-helper1 cells specific for M. leprae antigens are capable of activating macrophages and inducing destruction of M. leprae. The bacteria generally replicate very slowly under the pressure of this effective immune response and the disease progresses very slowly. Skin and peripheral nerves are damaged by the inflammatory responses that result from macrophage activation (Janeway et al., 2005).
Lepromatous leprosy, on the other hand, results from the failure to produce an effective cell-mediated immune response and predominance of humoral immune responses. Secretion of IL-4 and IL-6 lead to the T cell differentiation into the T-helper2 subtypes. B cells are activated to make neutralizing antibodies, predominantly of the IgM, IgA, and IgE classes. These antibodies are not able to reach intracellular bacteria and do not constitute an effective immune response. M. leprae are able to grow abundantly in macrophages and cause gross tissue destruction. Infection is widespread throughout the body, resulting in extensive damage to bones, cartilage, and nerves (Janeway et al., 2005).
Once an immune response has been established, it is not likely to switch from a humoral to a cell-mediated response or vice-versa because the two CD4 T-cell subsets act to negatively regulate each other. T-helper2 cells secrete IL-10 and TGF-β, which inhibit the activation and growth of T-helper1 cells. Conversely, T-helper1 cells secrete IFN-γ, which inhibits the proliferation of T-helper2 cells.
It is thought that certain subclasses of CD8 T-cells can respond to antigen by secreting cytokines typical of either T-helper1 or T-helper2 cells, and therefore determine which response will predominate. In the lepromatous form, CD8 T cells secrete IL-10 and TGF-β, inhibiting T-helper1 cells and leading to the default humoral response (Janeway et al., 2005). The study of these cells is critical to understanding the development of the disease in the tuberculoid or the lepromatous form and the potential development of treatment strategies aimed at redirecting the cytokine response.
The importance of cytokine production in disease progression was further demonstrated by examining the cytokine profiles in tuberculoid and lepromatous skin lesions. Cytokines resulting from macrophage activation, such as IL-1β, TNF-α, GM-CSF, TGF-β1, and IL-6, were much more plentiful in tuberculoid lesions than lepromatous lesions. Cytokines produced by T-cells, including IL-2, IFN-γ, and lymphotoxin, were likewise more abundant in tuberculoid lesions. IL-4, IL-5, and IL-10 cytokines where produced in lepromatous lesions but virtually absent in tuberculoid lesions (Yamamura et al., 1991). Consistent with the predominance of a humoral immune response, these cytokines induce B cells to produce neutralizing antibodies of the IgA, IgE, and IgM isotypes; IL-10 in addition serves as a potent suppressor of macrophage function.
The large quantity of TNF-α in tuberculoid lesions have been implicated in the gene expression of pro-apoptotic members of the Bcl-2 protein family, including Bad, Bax and Bak proteins. Induction of apoptosis in M. leprae-infected cells can decrease the bacterial load and make the environment of macrophages more hostile, limiting the replication of the bacteria (Hernandez et al., 2003). This programmed cell death is one way in which infection is contained in patients with tuberculoid leprosy.
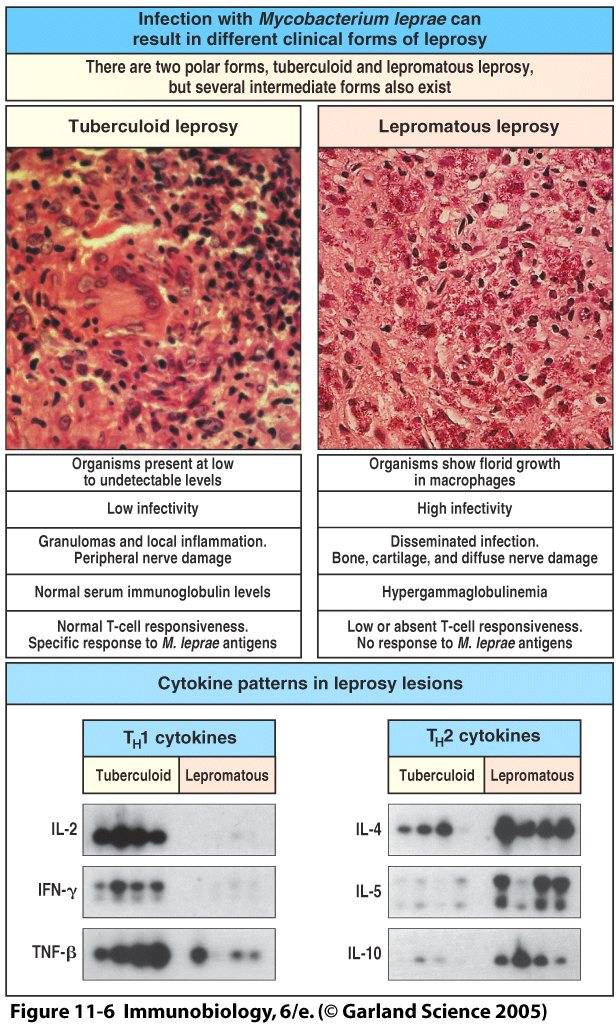
Fig. 2. Tuberculoid and lepromatous manifestations following M. leprae infection differ in their cytokine patterns, number of bacteria, infectivity, extent of damage, and containment by the immune response (Janeway et al., 2005).
Historically, there has been a lot of disagreement about whether genetics played a role in the development of leprosy. It is now understood that infection with M. leprae is the determining factor in the establishment of disease, but some genetic influence in its progression has been recognized. Specific HLA-DR alleles have been shown to play a role in mediating T-cell responses in conjunction with mycobacterial antigens. These alleles do not determine whether an individual will develop leprosy at all, but whether they will develop the tuberculoid or lepromatous form. The HLA-D region codes for class II antigens, which are expressed on the surface of macrophages and involved in antigen presentation to T helper or T suppressor cells (Fine, 1988). As we have already seen, the peptides presented influence T helper cell differentiation.
The current treatment strategy for Hansen’s disease is a multi-drug therapy approach. This strategy combines Rifampicin, Dapsone, and Clofazimine. Use of the three drugs in combination prevents the development of resistance. While monotherapy can result in drug resistance and therefore be a problematic treatment option, there have not been reports of resistance to multi-drug therapy (Ishii, 2003).
Rifampicin is a potent bactericidal agent against M. leprae and is administered once a month to prevent toxic effects of the drug. Bacterial multiplication is prevented at the time the drug is taken and over the next few days it kills over 99.9% of viable bacteria.
Dapsone was the first drug discovered to be effective against M. leprae in 1940 and was for many years the only drug used in the treatment of Hansen’s disease. Development of some resistant microbial strains made it necessary to use dapsone in conjunction with other drugs.
Clofazimine preferentially binds to bacterial DNA, thereby inhibiting mycobacterial proliferation and acting as a slow batericidal agent. It is to be taken on a daily basis.
Three more drugs have recently been shown to have bactericidal effects against M. leprae: ofloxacin, minocycline, and clarithromycin. However, due to cost-effectiveness and success using the traditional drugs, the World Health Organization’s multi-drug therapy recommendations have not changed.
It is generally assumed that paucibacilar Hansen’s disease can be cleared within six months of treatment, whereas multibacilar leprosy may take as long as twelve to twenty-four months for infection to clear. Relapse rates are extremely low.
The BCG (Bacille Camelette-Guerin) vaccine that is widely used in developing countries for protection against Mycobacterium tuberculosis has also been shown to have some protective qualities against leprosy. However, instituting a global BCG vaccination program against leprosy is not an economically feasible or effective plan (Ishii, 2003).
Other treatment strategies have been proposed involving the injection of cytokines to redirect a humoral immune response with predominantly T-helper2 cells to a cell-mediated one involving primarily T-helper1 cells. Lesions may be controlled or even cured more easily if a lepromatous lesion is transformed into a tuberculoid one (Janeway et al., 2005).
Bibliography
Centers for Disease Control and Prevention: National Center for Infectious Diseases/Division of Bacterial and Mycotic Diseases. “Hansen’s Disease (Leprosy).” Rev. Oct. 2005. http://www.cdc.gov/ncidod/dbmd/diseaseinfo/hansens_t.htm
Fine, P.E.M. “Implications of Genetics for the Epidemiology and Control of Leprosy.” Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. Vol. 321.1207, The Epidemiology and Exology of Infectious Disease Agents. 1988: 365-376.
Hernandez, M.O. et al. “Induction of Apoptosis in Monocytes by Mycobacterium leprae in vitro: a Possible Role for Tumor Necrosis Factor-α.” Immunology. Vol. 109, 2003: 156-164.
Ishii, Norihisa. “Recent Advances in the Treatment of Leprosy.” Dermatology Journal Online. Vol.9, No.2. 2003. http://dermatology.cdlib.org/92/reviews/leprosy/ishii.html
Janeway, C.A., Travers, P., Walport, M., Shlomcik, M. Immunobiology: The Immune System In Health and Disease. 6th ed. New York: Garland Science, 2005.
Mohanty, et al. “Leprosy Reactions: Humoral and Cellular Immune Responses to M. leprae, 65kDa, 28kDa, and 18kDa Antigens.” International Journal of Leprosy and Other Mycobacterial Diseases. Vol. 72, No. 2. 2002: 149-158.
Nardell, Edward A. and Willy F. Piessens. “Transmission of Tuberculosis” Tuberculosis: A Comprehensive International Approach. Eds. Lee B. Reichman and Earl S. Hershfield. 2 nd ed. New York: Marcel Dekker, Inc., 2000. 215-240.
Robbins, Stanley L and Ramzi S. Cotran. Pathological Basis of Disease. Ed. Vinay Kumar, Abul K. Abbas, and Nelson Fausto. 7 th ed. Philadelphia: Elsevier Saunders, 2005.
Schorey, Jeffrey, Michael C. Carroll, Eric J. Brown. “A Macrophage Invasion Mechanism of Pathogenic Mycobacteria.” Science, New Series, Vol.277.5229 1997: 1091-1093.
Sieling, P.A. et al. “CD1-Restricted T Cell Recognition of Mirobial Lipoglycan Antigens.” Science, New Series, Vol 269.5221, 1995: 227-230.
Sieling, Paul A. et al. “The Human CD1-Restricted T Cell Repetoire is Limited to Cross-Reactive Antigens: Implications for Host Responses Against Immunologically Related Pathogens.” The Journal of Immunology. Vol. 174, 2005: 2637-2644.
Wani, Junaid, Sabia Rashid, Muzaffar Sherwani, and A.R. Nasti. “Ocular Manifestations of Leprosy – A Clinical Study.” JK-Practitioner. Vol. 12(1): 14-17, 2005.
World Health Organization. “Leprosy fact sheet.” Rev. Oct. 2005. http://www.who.int/mediacentre/factsheets/fs101/en/
Yamamura, Masahiro, et al. “Defining Protective Responses to Pathogens: Cytokine Profiles in Leprosy Lesions.” Science, New Series, Vol.254.5029 1991: 277-279.