DISCUSSION
The GA1 phenotype has the potential of providing new insights into the mechanism used by Chlamydomonas for cell-cell interactions. These insights have inspired research aimed at identifying the molecular components of signalling pathways downstream of adhesion molecules and the mechanisms by which these molecules interact with other intracellular signalling systems to generate complex cell behaviors. The present study suggests that the GA1 gene is not an endosomal H+-ATPase. Drugs that specifically block H+-ATPases and increase intracellular pH levels were unable to mimic the GA1 mutant phenotype in wild-type cells. Therefore, the model that GA1 is an essential gene necessary for the proper glycosylation of agglutinin was not supported. However, sequence analysis of GA1 cDNA indicated that the gene is likely to encode an H+-ATPase of some sort (Table 4). The GA1 amino acid sequence is very similar to a "probable" E1-E2 cation-ATPase of Leishmania donovani, a unicellular flagellated parasite and to a known H+-transporting ATPase of Heterosigma akashiwo, a unicellular flagellated plankton algae (Appendix A). Future research should focus on these similarities (i.e. compare mating processes, analyze flagellar functions, and assess cell-cell interactions).
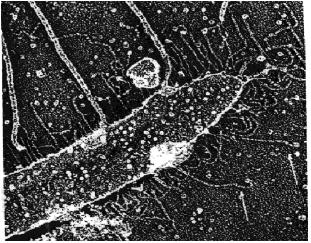
Until the whole gene has been cloned, we cannot rule out that the gene might encode the mt-agglutinin. Thus, further research needs to address this possibility. If funding and equipment are available then deep freeze etch imaging could be used to highlight the mt- agglutinins on the flagellar surfaces. Images of wild type mt- cells would have to be compared to images of GA1 cells to see if agglutinins are present in one strain and absent in the mutant strain (Figure 15).
Figure 15. Example of a deep freeze etch image of a wild type mt- gamete flagellum. The arrows are pointing to molecules with the morphology of mt- agglutinins (i.e. they are hooked at one end (Figure 1)). If GA1 encodes the mt- agglutinin then these molecules would be absent in a deep freeze etch image of GA1 flagellum. (Photo by Dr. John Heuser; Adair, 1985).
The second half of the putative GA1 gene still needs to be cloned. Subcloning methods like the ones performed in this study are currently being conducted to subclone the Bam HIfragment (Figure 16). Once this fragment is cloned then it can be ligated to the Sal I fragment and used in transformation experiments to complement theGA1 mutation.
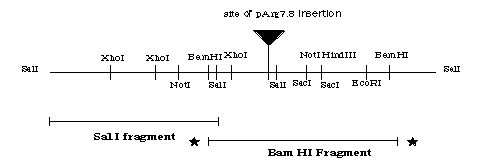
Figure 16. Restriction enzyme map of the putative GA1 gene indicating the Sal I fragment that has been subcloned and the Bam HI fragment that needs to be sublconed.
Once the putative GA1 gene has been cloned, the specific fragment that encodes the actual gene can be isolated by making deletions in the gene and then performing transformation experiments to complement the GA1 mutation. This process would be done successively until a fragment from the putative GA1 gene was discovered that complements the behavioral mutation.
C. reinhardtii, namely GA1, has tremendous potential to provide new approaches for elucidating important problems in signal transduction. The mating system provides an excellent opportunity for the molecular dissection of coupling extracellular matrix and the redistribution of integral membrane proteins to signalling pathways (Quarmby and Hartzell, 1994). In general, there is a poor understanding of signal transductions and their relations with cell-cell interactions; fortunately, the use of C. reinhardtii provides molecular and cell biologists with the opportunity to address such an important question.
I must express my deepest appreciation for Dr. A. Malcolm Campbell for his unending guidance and support, timeless patience, and strong commitment to the education of others. I am so thankful for having had the opportunity to work "one on one" with such an excellent teacher. He has truly been a positive influence in my life and I look to him not only as my professor but also as my good friend. I am grateful for Dr. John H. Williamson for his assistance in the lab as well as his encouragement throughout this project. Thanks to Dr. Virginia Armbrust who provided me with an interesting research topic. I would also like to thank Dr. Donald L. Kimmel, Dr. Jeremiah L. Putnam, Dr. Patricia A. Peroni, Mr. James W. Lowery, and the rest of the Biology faculty and staff who repeatedly let me in and out of rooms, answered questions, and provided me with that extra "pat on the back" when I needed it the most. I would also like to recognize Davidson College for providing me with the funds and the place that allowed me to have a very meaningful and memorable senior year experience. Lastly, I want to extend my love and gratitude to my friends and family for their generosity, support, and love.