This web page was produced as an assignment for an undergraduate course at Davidson College.
What DNA Ligase Orthologs Can Tell Us
Orthologs:
Orthologs are genes that share a common ancestor (Campbell, 2005). Gene orthologs can tell us much about a protein by examining highly conserved areas in the gene throughout the evolution of species. Information concerning various phenotypes associated with the protein, drug binding sites, and the evolution of various mutations, among other things, can be determined through the examination of gene orthologs.
Using Ensembl and NCBI, eight orthologs of DNA Ligase were obtained, including Arabidopsis thaliana, Drosophila melanogaster (fruit flies), C. elegans, E. coli, Mus musculus (mice), Chlorella virus, Homo sapians (humans), and Schizosaccharomyces pombe (fission yeast). To view the amino acid sequences of each ortholog, click on the links above. Each species listed above uses at least three forms of DNA ligase. Before the conservation sequences of orthologs are discussed, it is necessary to understand the perceived function of each enzyme.
DNA Ligase I
This variety of DNA ligase functions in DNA replication and also aids in the exonuclease repair mechanism. Mutations in the LIG1 gene cause an increased mutation rate when exposed to mutagens as well as a decrease in immune response (NCBI). It has been found that DNA ligase I is necessary for embryonic development, but it is not essential to the replication of DNA (Bentley et al., 1996).
DNA ligase III
DNA ligase III is thought to be involved in DNA repair and recombination. Two isoforms of DNA ligase III exist, alpha and beta. These isoforms are a result of alternative splicing of the LIG3 gene. Each is involved in a different role in organisms due to differences in the C termini of the protein sequences. The beta isoform, for example, is present only in the testis and is thought to play a pivotal role in recombination during spermatogenesis. The alpha isoform, on the other hand, is not tissue specific and is thought to play a role in DNA repair (Entrez Gene).
DNA ligase IV
This variety of DNA ligase alone has been found to dictate the actual ligation of damaged DNA strands in human cell lines(Grawonder et al., 1998). Mutations in DNA ligase IV have been indicated in cases of LIG4 Syndrome. This syndrome results from homozygocity in a three amino acid substitution on the LIG4 gene. Only one of the amino acid substitutions takes place in the highly conserved motif I, which contains the adenylation site of DNA ligase. The other two mutations are thought to aggravate the phenotype caused by the first mutation. A characteristic phenotype of LIG4 Syndrome includes retardation in physical and mental development of children, microcephaly at birth, pancytopenia, as well as a large presence of plantar warts (OMIM). However, DNA ligase IV has also been shown to aid in the prevention of Multiple Myeloma. A polymorphism in the LIG4 gene, resulting in a C-T substitution, is known to reduce a patient's risk of Multiple Myeloma by a factor of two. It is possible that this substitution alters the gene dosage in a favorable manner (OMIM).
*The locus of each isoform of DNA ligase in selected orthologs can be found at the following links:
To view the location of Drosophila DNA Ligase I, III, and IV, visit Ensembl.
To view the location of murine DNA Ligase I, III, and IV, visit Ensembl.
To view the location of human DNA Ligase I, III, and IV, visit Ensembl.
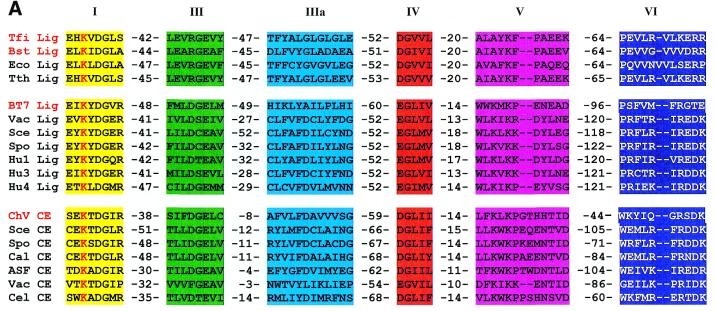
Although DNA ligases vary in their size, amino acid sequence, and properties, there are six motifs that are thought to make up the active sites of most ATP-dependent DNA ligases. Five of these six motifs also occur in NAD+-dependent DNA ligases (Aidan, 2000). Each of the motifs depicted in Figure 1 have been observed to contain sequences that have historically aided in catalysis, indicating further that these regions of the enzyme cluster at the active site. In motif I, the sequence Lys-(Tyr/Ala)-Asp-Gly-(Saa)-(Arg) is conserved in the active site. A similar sequence is also preserved in the NAD+-dependent ligases. The pivotal amino acid, it seems is the lysine residue in motif I. It is conserved in both ATP-dependent and NAD+-dependent DNA ligases, as well as in RNA capping enzymes. Therefore, the conclusion drawn by Aidan, that leucine is the site of adenylation, is supported by this data.
It is possible that when these conserved domains are fully understood in both ATP-dependent and NAD+-dependent DNA ligases, this information could aid scientists in synthesizing specific bacterial ligase inhibitors. Thus, further knowledge of the DNA ligase active site could lead to new antibacterial technology (Aidan, 2000).
Figure 1 Conserved motifs in DNA ligases as well as RNA capping enzymes. Motifs are assigned the numbering I, II, III, IV, V, and VI and represent both NAD+-dependent and ATP-dependent DNA ligases. Ligases encoded by T.filiformis (Tfi), B.stearothermophilus (Bst), E.coli (Eco) , and T.thermophilus (Tth) are NAD+-dependent DNA ligases. Ligases encoded by bacteriophage T7 (BT7), vaccinia virus (Vac), Saccharomyces cerevisiae (Sce), Schizosaccharomyces pombe (Spo), and human ligases I (Hu1), III (Hu3), and IV (Hu4) are ATP-dependent DNA ligases. Below the ATP-dependent ligases are RNA capping enzymes denoted by the suffix CE. These include Chlorella virus PBCV-1 (ChV), S.cervisiae, S.pombe, Candida albicans (Cal), African swine fever virus (ASF), vaccinia virus, and Caenorhabditis elegans (Cel). The dashed numbers separating amino acid sequences indicate the number of amino acids separating conserved motifs. The lysine residue of the active site is shown here in red. This image is courtesy of Aidan, 2000 ,Figure 3A,(permission pending).
Besides the indicated motifs, DNA ligases differ immensely in their amino acid sequences across species. This is due largely to and increased specificity through additional ligase domains. Even with amino acid sequence diversity, however, the protein conformation through tertiary structure is preserved through similar hydrophilic and hydrophobic regions (Tomkinson et al., 1991). These regions are conserved across species to a great degree. To view the conservation of hydrophobic and hydrophilic regions in DNA ligase orthologs, click here.
Works Cited:
Aidan, J., Suh, S. 2000. Structural and mechanistic conservation in DNA ligases. Nucleic Acids Research 28(21): 4051-58.
Bentley D., Selfridge J., Millar JK., Samuel K., Hole N., Ansell JD., Melton DW. 1996. DNA ligase I is required for fetal liver erythropoiesis but is not essential for mammalian cell viability [abstract]. Nature Genetics 13(4): 489-91.
Campbell, M. 2005. Standards for Molecular Web Page Assignments. http://bio.davidson.edu/courses/Molbio/standardsHP.html#anchor78181983. Accessed 2005 March 6.
Ensembl. Ensembl Genome Browser: DNA ligase. Accessed 2005 March 8.
Grawonder, U., Zimmer, D., Fugmann, S., Schwarz, K., Lieber, MR. 1998. DNA Ligase IV is essential for V(D)J recombination and DNA double-strand break repair in human precursor lymphocytes [abstract]. Mol Cell 2(4): 477-84.
NCBI. BLAST: DNA Ligase. Accessed 2005 March 6.
NCBI. Entrez. Entrez Gene: LIG3. Accessed 2005 March 6.
NCBI. [OMIM]Online Mendelian Inheritance in Man. Johns Hopkins University. Accessed 2005, March 9.
[PDB]Protein Data Bank. Structural Explorer-1P8L. Accessed 2005 March 8.
Tomkinson, A., Totty, N., Ginsburg, M., Lindahl, T. 1991. Location of the active sit for enzyme-adenylate formation in DNA ligases. Proc. Natl Acad. Sci. USA, 88: 400-404.
Davidson College Biology Home Page
© Copyright 2005 Department of Biology, Davidson College, Davidson, NC 28036
If you have any questions, comments, or suggestions concerning this page, please contact sadurnbaugh@davidson.edu