*This website was produced as an assignment for an undergratuate course at Davidson College.*
Induced Pluripotent Stem Cells Review Paper
Overview of Induced Pluripotent Stem Cells
Embryonic stem (ES) cells are unspecialized cells capable of renewing themselves through cell division. Being pluripotent, they have the remarkable potential to develop into many different cell types with specialized function, depending on the environment. Thus, ES cells are readily becoming a major topic of exploration in the medical field as they could be the answer to the possible treatment of a multitude of human diseases such as diabetes and heart disease. ES cells can be grown in cell culture, induced to differentiate into a certain tissue type, and then inserted into a patient with the disease. However, ethical debates still surface over ES cells regarding the idea of destroying human embryos. In their paper, “Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors,” Takahashi and Yamanaka examine whether or not somatic cells could revert from their specialized, tissue-specific function to a state of pluripotency, such as that in ES cells. These somatic cells that have been genetically reprogrammed to an ES cell-like state are known as induced pluripotent stem cells, or iPS cells. Medically, the iPS cells are desirable because patient’s own cells could be reverted into the pluripotent stage to produce healthy tissue, avoiding the unfortunate case of a patient’s immune system rejecting foreign tissue produced from ES cells. Several transcription factors, such as Oct3/4, Sox2, and Nanog, have been determined candidates for maintaining the unique pluripotent characteristics of stem cells. Takahashi and Yamanaka attempt to demonstrate that pluripotency can be induced in terminally differentiated somatic cells by the combination of four transcription factors, Oct3/4, Sox2, c-Myc, and Klf4.
Results
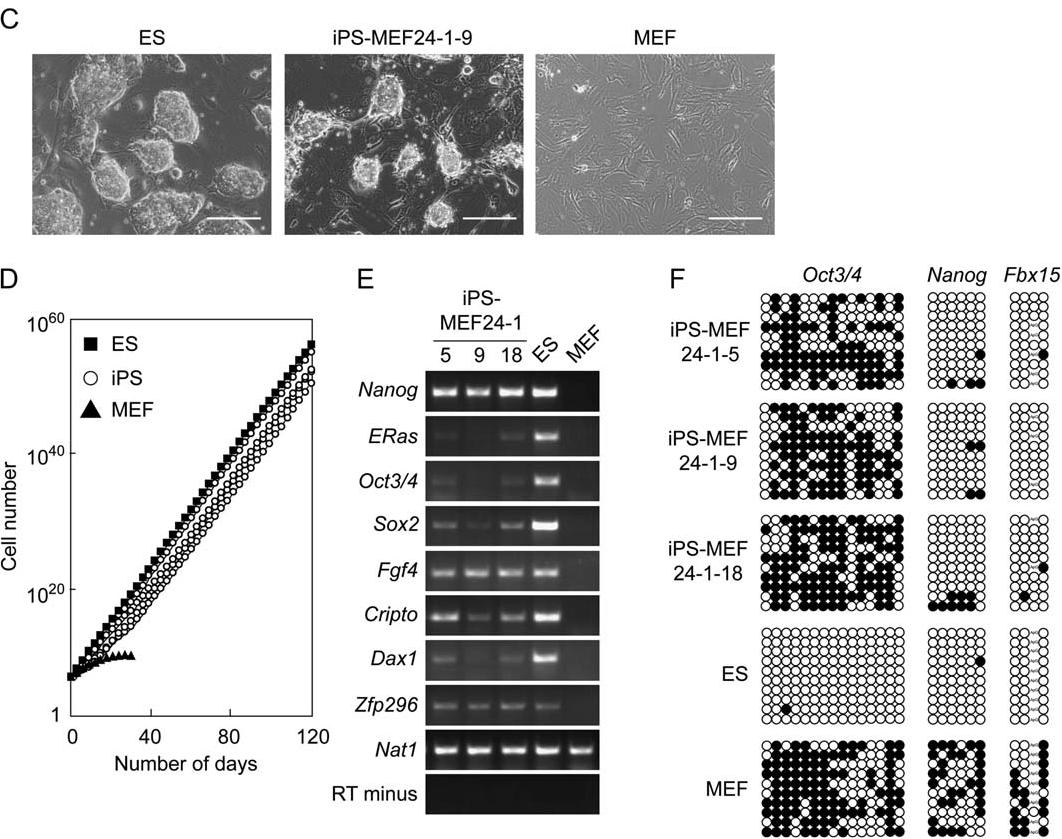
To test for the potential induction of pluripotency, Takahashi and Yamanaka began by selecting 24 genes previously suspected to contain factors important for maintaining the defining properties of ES cells. A fusion of the β-galactosidase and neomycin resistance genes (βgeo) was inserted by homologous recombination behind the promoter of the mouse Fbx15 gene and selected for G418 resistance (Figure 1A). ES cells of Fbx15 βgeo/βgeo genotype could tolerate much higher concentrations of G418 than somatic cells of the same genotype. Each of the 24 candidate genes were retrovirally transducted into mouse embryonic fibroblasts (MEFs) from Fbx15 embryos homozygous for βgeo and selected for G418 resistance. Because Fbx15 is a gene that is only expressed in ES cells and early embryos and plays a role in the maintenance of pluripotency, cells expressing Fbx15 and tolerant to normal amounts of G418 after the addition of a combination of the 24 gene factors would thus be similar to ES cells. The transduction of a single factor into the MEFs did not produce any drug-resistant colonies grown on STO feeder cells, suggesting that more than one transcription factor is needed to turn on the Fbx15 gene for pluripotency. In fact, when all 24 of the candidate factors were transduced into the fibroblast together, 22 G418-resistant colonies grew (Figure 1B). 5 selected clones displayed physical characteristics similar to ES cells, such as a round shape, large nucleoli, and little cytoplasm. Figure 1C gives a morphological comparison between ES, iPS-MEF24-1-9 (induced pluripotent stem cells created from MEF with all 24 candidate factors), and MEF. The similarities between ES and iPS-MEF24-1-9 are striking, even though iPS-MEF24 cells started as simple MEF cells. In addition, proliferation of cells was remarkably similar between the ES and iPS cells (Figure 1D). Both of these cells showed continuous linear growth characteristic of stem cells for over 120 days, while MEF proliferation horizontally leveled off in less than 40 days. Reverse transcriptase PCR (RT-PCR) analysis shows that genes normally expressed within ES cells, such as Oct3/4, Nanog, ERas, Cripto, Dax1, Zfp296, and Fgf4, were also expressed in iPS-MEF24 clones (Figure 1E). All iPS-MEF24 clones clearly expressed Nanog. Yet, not all genes expressed in one ES cell were also expressed in an individual iPS clone. This suggests that not all ES cell marker genes are required for inducing pluripotent characteristics. Supporting this claim is Figure 1F. Bisulfite genomic sequencing shows that the promoters of Fbx15 and Nanog were demethylated—an indicator of expression—while the Oct3/4 promoter remained methylated in iPS-MEF24 cells.
Figure 1C, D, E: iPS-MEF24 clones show striking similarities to ES cells in the morphological appearance, growth rate, and cell marker expression.The demethylation of Nanog and Fbx15 promoters is also similar to the type of methylation in ES cells.
Takahashi and Yamanaka needed to narrow down the candidate factors to the necessary factors required to induce reversion to pluripotent cells. They did this by removing individual factors and noting whether or not colonies formed. Ten factors (known as 3, 4, 5, 11, 14, 15, 18, 20, 21, and 22) were found to have no colony formation or very minimal colony formation (Figure 2A). Interestingly, when combinations of just these 10 factors were transduced into the MEF cells, more iPS colonies resulted than when all 24 factors were incorporated (Figure 2B). From these 10 factors, each factor was then individually removed to test for colony formation. Four factors—Oct3/4, Klf4, Sox2, and c-Myc—were found to initiate substantial G418-resistant colony formation, suggesting that these factors are very important players in the generation of iPS cells from MEFs. Takahashi and Yamanaka attempted to narrow the number of crucial factors to two or three factors, but results were not particularly successful as shown in Figure 2C. No two factor combinations could promote colony formation. Combinations of three genes, including the mixture of Oct3/4, Sox2, and c-Myc and the mixture of Klf4, Sox2, and c-Myc, could not produce colonies that proliferated in culture. The combination of Oct3/4, Klf4, and Sox2 produced 36 G418-resistant colonies, but the morphology did not mirror that of ES cells. Lastly, the mixture of factors lacking Sox2 produced 54 colonies in which the clones could be maintained in culture. These clones, iPS-MEF3, were compared to the appearance of the iPS-MEF4 and iPS-MEF10 cells (cell lines created by the transduction of the four crucial transcription factors or the initially determined 10 factors) in Figure 2D. Clearly, the iPS-MEF4 and iPS-MEF10 cells share similar morphology, while iPS-MEF3 cells show jagged, rough surfaces. So although the Fbx15 locus can be expressed in the iPS-MEF3 cells, the four transcription factors are required for ES cell-like morphology. Thus, the transcription factors Oct3/4, Klf4, Sox2, and c-Myc were determined necessary and sufficient for the induction of iPS cells from MEFs.
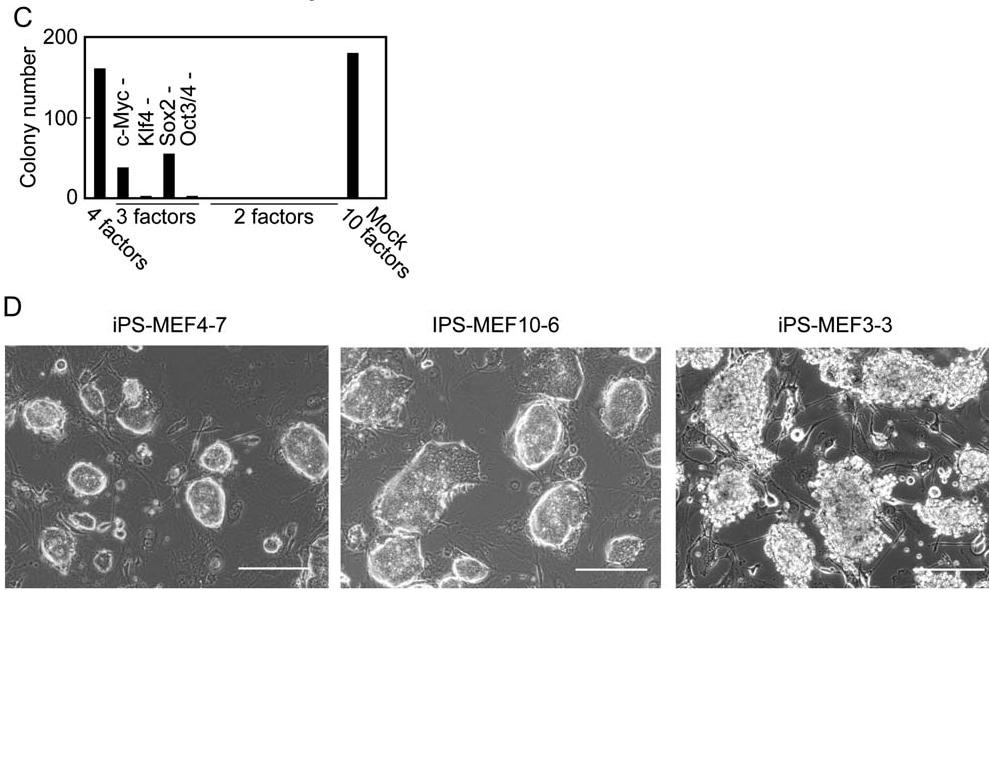
Figure 2:C) The combination of the four critical transcription factors of OCt3/4, Sox2, c-Myc, and Klf4 is necessary for large iPS-MEF colony growth. 3-gene combinations in iPS-MEF cells still can produce colonies, but to a lesser extent. D) There are clear morphological differences between the iPS-MEF colonies that contain the four transcription factors and the iPS-MEF3 cells that do not.
Takahashi and Yamanaka turned to RT-PCR to observe the expression of ES cell marker genes in iPS cells. Figure 3A looks at iPS-MEF3 (-Sox2), iPS-MEF4 (Oct3/4, Klf4, Sox2, and c-Myc), and iPS-MEF10 (all 10 factors) as well as control ES and MEF cells. In general, iPS cells were found to express similar genes to the ES cells, but were not identical. iPs-MEF4 and iPS-MEF10 cells expressed every gene except for Ecat1. iPS-MEF3 cells failed to express marker genes including Ecat1, Esg1, Oct3/4, and Sox2, suggesting that iPS-MEF3 cells are significantly different from the cell lines containing the Sox2 transcription factor. This lack of upregulation could also be a reason why iPS-MEF3 cells are not pluripotent. This substantial difference was further confirmed by more evidence in the DNA microarray approach in Figure 4. Microarray gene expression results revealed that iPS-MEF3 did not successfully upregulate Dppa3, Dppa4, Dppa5, Nanog, Sox2, Esrrb, and Rex1 as did ES, iPS-MEF4, and iPS-MEF10 cells. Statistical results from Pearson correlation analysis explained that iPS cells are more closely related to ES cells than fibroblasts. However, Takahashi and Yamanaka once again did not shy from admitting that iPS-MEF cells were not identical to ES cells. Microarray results showed that only ES cells upregulated Dnmt3a, Dnmt3b, Dnmt3l, Utf1, Tcl1, and the gene for the LIF receptor. Takahashi and Yamanaka chose two cell clones that appeared to be most similar to ES cells—iPS-MEF4-7 and iPS-MEF10-6—and examined them further. To analyze methylation and acetylation of promoters, chromatin immunoprecipitation was utilized for the promoters of Oct3/4 and Nanog (Figure 3B). Results showed that the promoters remained partially methylated in iPS cells, illustrating yet another difference between iPS cells and ES cells (Figure 3C).
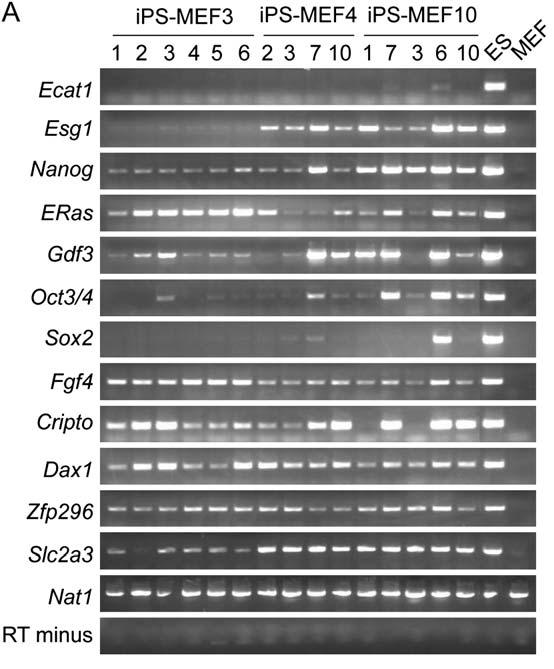
Figure 3A: RT-PCR shows that, except for Ecat1, iPS-MEF10 and iPS-MEF4 transcribed most of the cell marker genes in ES cells.iPS-MEF3 genes failed to express several marker genes. iPS-MEF cells can be closely identified with ES cells, but are not identical.
Though Takahashi and Yamanaka had shown that iPS-MEF cells expressed many of the cell markers present in ES cells, there was no conclusive evidence pointing to the pluripotency of these iPS cells. Thus, they made the move from cells growing on plates to injecting the cells into mice. Once injected into nude mice, the iPS cells caused the formation of teratomas, or tumors with cells from all three germ layers. The majority, but not all, of the iPS-MEF10 and iPS-MEF4 clones demonstrated pluripotency. Of the colonies injected into the mice, iPS-MEF4-2 and -7 as well as iPS-MEF10-3 and -6 gave rise to tumors of all three germ layers, containing nervous, connective, and epithelial tissue as revealed through immunostaining (Figure 5A and 5B). Figure 5C reveals that embryoid bodies, or accumulations of cells in tissue culture dishes, from iPS-MEF3 were composed of undifferentiated cells. In contrast, iPS-MEF4-7 and iPS-MEF10-6 cells initiated differentiation into three germ layers in the embryoid bodies, as shown by the different staining in Figure 5D.
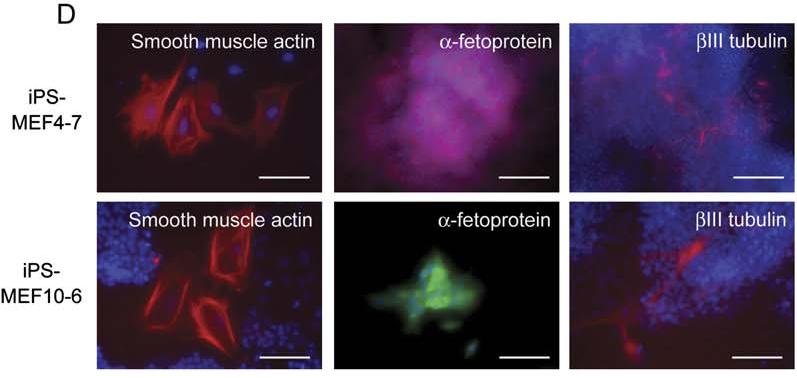
Figure 5D: iPS-MEF4-7 and iPS-MEF10-6 cells displayed pluripotent characteristics by differentiating into all three germ layers. Immunostaining detected cells positive for smooth muscle actin (mesoderm marker), α-fetoprotein (endoderm marker), and βIII tubulin (ectoderm marker).
After showing that iPS-MEF cells were pluripotent, Takahashi and Yamanaka introduced the four selected transcription factors—Oct3/4, Klf4, Sox2, and c-Myc—into the tail-tip fibroblasts (TTFs) of four 7-week-old male Fbx15βgeo/βgeo mice and a 12-week-old female Fbx15βgeo/βgeo mouse that also expressed green fluorescent protein (GFP). G418-resistant colonies formed and iPS-TTFgfp4 cells were established that were physically indistinguishable from ES cells, suggesting once again that the four factors are necessary and sufficient to produce iPS cells (Figure 6A). In addition, the iPS-TTFgfp4 cells were shown through RT-PCR to express most ES cell marker genes at high levels and a few at lower levels (Figure 6B). iPS-TTFgfp4-3 and iPS-TTFgfp4-7 expressed all genes except for Ecat1, similarly to what the authors claimed iPS-MEF10-6 and iPS-MEF4-7 expressed. Just as before, the results revealed that iPS-TTF cells are similar, but not identical to ES cells. Proof of pluripotency resulted after 2 iPS-TTF4 and 6 iPS-TTFgfp4 clones were transplanted into mice and tumors consisting of cells from all three germ layers formed. The clones iPS-TTFgfp4-3 and iPS-TTFgfp4-7 were introduced into blastocysts and subsequently contributed to the development of 18 iPS-TTFgfp4-3 and 22 iPS-TTFgfp4-7, GFP-positive embryos (Figure 6C). In the embryos, iPS-TTFgfp4 cells could differentiate into all three germ layers—the hallmark of pluripotency—as illustrated by histological slides in Figure 6D. The authors showed that, like ES cells, iPS can to contribute to development.
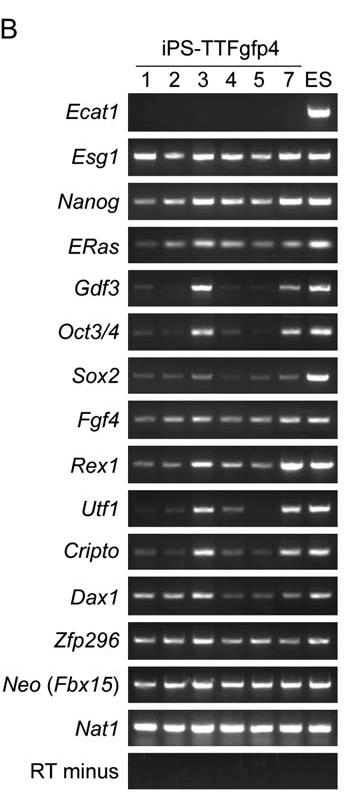
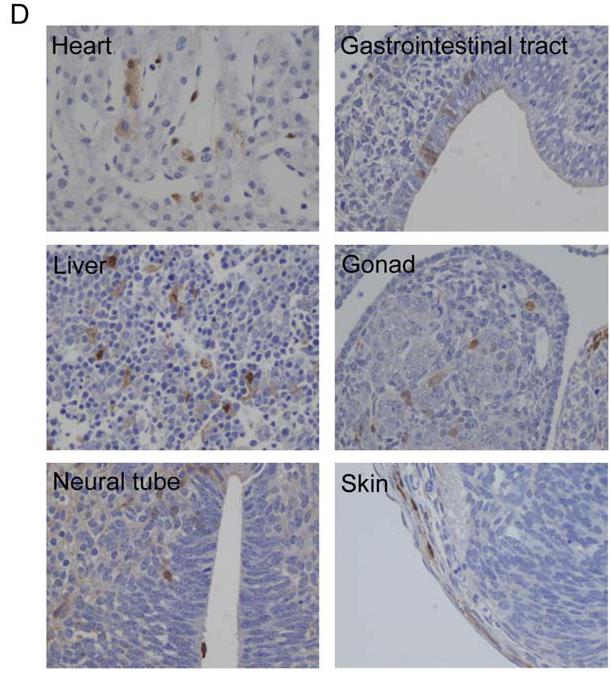
Figure 6A: iPS-TTFgfp4 cells expressed many of the cell marker genes found expressed in ES cells. D) Brown staining reveals the presence of GFP-positive iPS-TTFgfp4 cells. These cells are expressed in all three germ layers.
The last figure in Takahashi and Yamanaka’s paper summarizes the results of the created iPS cells, combining data for iPS-TT4gfp4 and iPS-MEF cell clones. Transduction of iPS cells into fibroblasts did not lead to overexpression of protein product within the cells. Based from the loading control β actin in the Western Blot in Figure 7A, the same amounts of a specific protein were detected in the ES cells and the iPS cells. Translation of Nanog and ERas expression was visibly greater in ES cells than iPS cells. However, iPS-MEF10-6 cells did translate these two proteins because, although neither were one of the four crucial factors, these two factor transcripts were initially transduced into the iPS-MEF10 clones. Figure 7B discusses the idea that undifferentiated iPS-MEF 4-7, iPS-MEF10-6, and iPS-TTFgfp4-3 cells as well as ES cells show higher Oct3/4, Sox2, and Nanog RNA and protein expression levels than differentiated cells. These specific RNAs and proteins are not expressed at all in MEF and TTF cells since they are specific to ES and ES-like cells. Southern Blot analysis with a Klf4 probe in Figure 7C reveals that over 20 transgenes from the retrovirus was incorporated into the DNA of every iPS clone. Thus, the integration of some of these transgenes could contribute to gene silencing or other expression consequences. Generally, the iPS-TTFgfp4 clones had normal karyotypes, suggesting that iPS cell incorporation does not affect chromosomal structure or content (Figure 7D). However, minor chromosomal alterations that cannot be detected by karyotype must not be dismissed. When iPS cells were cultured in environments without feeder cells, the iPS cells could not remain undifferentiated, even in the presence of Leukemia inhibitory factor (LIF) which tends to restrict cells from moving towards differentiation (Williams et al, 1988). While ES cells aggregated and tended to differentiate under the presence of LIF, iPS-MEF4-7 and iPS-TTFgfp4-3 cells remained undifferentiated and disassociated (Figure 7E).
Critique
Overall, Takahashi and Yamanaka presented convincing evidence that pluripotent stem cells with the morphology, growth properties, and cell marker expression of ES cells can be induced from mouse embryonic or adult fibroblasts after the introduction of four transcription factors—Oct3/4, Sox2, c-Myc, and Klf4. The experimental results in general support the authors' assertions. Figures throughout the paper were detailed and descriptive, and in most instances, more than one test was done to verify a claim, such as the several images in Figure 1 that show iPS cell similarity to ES cells. RT-PCR figures were usually clear, and the authors consistently applied both positive and negative controls to the figures. Every RT-PCR figure, except for Figure 6B, also had lanes showing ES and MEF expression of transcription factors, making comparisons between the transduced cells and ES or MEF easily obtained. Figure 6B should have had a control lane dedicated to TTF cell expression. Since the TTF cells are different from the MEF cells, we cannot assume that there is the same negative expression of ES cell markers. Microscopic images of morphology and histology were well prepared and showed a clear similarity in physical appearance between ES and iPS cells as well as obvious differentiation in the three germ layers.
I thought some figures did lack complete thouroughness. Figure 1E is difficult to interpret due to the level of expression for each clone. The authors seem to overinterpret the brightness of bands. Clearly, the iPS-MEF24 clones express the same levels of Nanog, Fgf4, Cripto, and Zfp296 as ES due to the similarity in brightness. However, there are visible differences in expression and band brightness between the iPS clones and ES with Eras, Oct3/4, Sox2, and Dax1. Though the iPS cells more closely assume the characteristic expression patterns of ES cells than MEF, I do not know if this RT-PCR could stand alone in convincing a reader that iPS-MEF cells followed the same expression patterns as ES cells. In Figure 2D, the authors show that the 3-gene combination lacking Sox2 resulted in cells with morphology not similar to ES cells. However, they neglected to show an image of the –c-Myc cells, even though they mentioned that these iPS cells did not resemble ES cells either. It is not mentioned whether or not the authors continued their observations of –c-Myc iPS cells, but it is a bit troubling that only –Sox2 cell information was included in the figure. In addition, the authors do not continue looking at –c-Myc cell lines in the rest of the paper. In Figure 3A, expression of marker genes also varied between different clones within the same cell lines, making the diagram difficult to interpret. The authors admitted that expression of several marker genes differed between clones, for example the higher expression of Oct3/4 in iPS-MEF4-7, iPS-MEF10-6, and iPS-MEF10-7 clones than in others. Yet, at the same time, there was no mention of the reason behind the lack of a band for Sox2 in many of the iPS-MEF4 and iPS-MEF10 clone lanes. In this figure, the authors seem to skim over results that do not support their claim of the similarity between iPS-MEF and ES cells. I believe the results have made it evident that there exist significant differences in the assortment of proteins and levels of expression of each cell type, and the authors have not fully addressed this issue. Figure 3D lacked a control figure showing cells negative for alkaline phosphatase and SSEA-1. Thus, this figure could lend little to my interpretation of the data and conclusions because I did not know what I needed to be looking for. Lastly, the images of Figure 5 are informative, but it would have been helpful to have control images of normal mouse tissue and normal ES cell activity. Though Takahashi and Yamanaka are likely catering their article to an audience of scientists who know what healthy, normal mouse tissue looks like, I believe it would have made their argument stronger to see the differentiated iPS-MEF cell-derived tissue in comparison with the normal germ layers.
I also think highly of the authors for admitting that their findings had shortcomings, and in no way had they perfected the science behind iPS cells. Takahashi and Yamanaka made it clear that though iPS cells shared many of the same characteristics of ES cells, they were not identical to ES cells and further work must be done on iPS cells to discover their origin and the role the four critical factors play in the reversion to pluripotency.
Future Experiments
Generation of stem cells using no embryonic material opens up impressive opportunities in the research world. From the medical perspective, an important future experiment would be to determine the success of iPS cells in human tissues. First, the genes controlling ES cell expression and maintenance of pluripotence would have to be determined. This could be accomplished through a microarray technique looking at the genes upregulated in ES cells in humans. Retrovirus transduction of the candidate transcription factors—such as those that behave like Oct3/4, Sox2, Klf4, and c-Myc genes—using a construct similar to the Fbx15βgeo/βgeo into human somatic cells could potentially lead to the formation of iPS cells in culture. Confirmation of ES cell gene expression in iPS cells could be determined with the RT-PCR method. If functional and pluripotent, thse iPS cells could have the potential to make healthy tissue in diseased patients. After hypothesizing this future experiment, a quick search on the internet showed me that Takahashi and Yamanaka in 2007 were able to make iPS cells in human fibroblasts (Takahashi et al., 2007). However, a complication to the incorporation of iPS cells into human somatic cells is the c-Myc factor. Takahasi and Yamanaka suggest that this factor may not be suitable for clinical applications. c-Myc is a proto-oncogene that encodes a transcription factor essential for cell growth and proliferation (Dominguez-Sola et al, 2007). It is responsible for tumorigenesis, and thus could induce tumor formation in humans--a problem that could be brought about by incorporating c-Myc in iPS cells.
Takahashi and Yamanaka were unable to create iPS cells identical to ES cells. The microarray showed many genes that were upregulated in the ES cells that were not upregulated in iPS cells such as Dnmt3a, Dnmt3b, Dnmt3l, Utf1, Tcl1, and the gene for the LIF receptor. What is important about these genes and why are they not activated in iPS cells? RT-PCR of these genes’ expression in iPS cells should be conducted to see whether or not iPS can activate these transcription factors. In addition, analysis of these genes in ES cells could show their function, and possibly why ES cells are unique from iPS cells. Immunostaining of these proteins in fibroblasts could show location. Sequencing these genes and subsequently individually knocking out one of the genes could give information regarding function. If it is found that one of these transcription factors is actually important to ES cell function, that gene could be incorporated into the Fbx15 βgeo/βgeo construct to develop new iPS cells.
In their paper, Takahashi and Yamanaka discussed that they were not sure why the four transcription factors—Oct3/4, Sox2, Klf4, and c-Myc—that they found were sufficient for iPS cell induction. As a result, a question to be asked and further explored is why these four factors? Other research has argued that factors such as Nanog and Lin28 are also necessary for iPS generation (Thompson et al, 2007). In addition, the figures throughout the paper suggest that other factors are still being expressed in iPS cells at equal or sometimes even greater amounts than the four “crucial” factors. I think it would be beneficial to continue looking into more transcription factors as relevant to the generation of iPS cells. In particular, Ecat, Fgf4, and Cripto should be looked at more closely. To determine whether or not these factors play a role in the reversion to pluripotency, a Western Blot showing protein amounts in iPS cells should be run. If these additional factors are important in iPS generation, then bands would show equal widths to those of ES cell proteins.
References
Dominguez-Sola D, Ying CY, Grandori C, Ruggiero L, Chen B, Li M, Galloway DA, Gu W, Gautier J, Dalla-Favera R. 2007. Non-transcriptional control of DNA replication by c-Myc. Nature 448 (7152): 445–51.
Stem Cell Information [World Wide Web site]. Bethesda, MD: National Institutes of Health, U.S. Department of Health and Human Services, [cited ] Available at <http://stemcells.nih.gov/info/basics/basics3>
Takahashi K, Yamanaka S. 2006. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126: 663-676.
Williams RL, Hilton DJ, Pease S, Willson TA, Stewart CL, Gearing DP, Wagner EF, Metcalf D, Nicola NA, Gough NM. (1988). Myeloid leukaemia inhibitory factor maintains the developmental potential of embryonic stem cells. Nature 336: 684 - 687.
Yu J, Vodyanik MA, Smuga-Otto K, Antosiewicz-Bourget J, Frane JL, Tian S, Nie J, Jonsdottir GA, Ruotti V, Stewart R, Slukvin II, Thomson JA. 2007. Induced pluripotent stem cell lines derived from human somatic cells. Science 318: 1865.
Please direct questions or comments to Sarah Little