*This website was produced as an assignment for an undergraduate course at Davidson College.*
Basic Question
In the paper entitled “Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors,” Takahashi and Yamanaka describe the reversion of differentiated fibroblast cells to pluripotent stem cells with the addition of four proteins: Oct3/4, Sox2, c-Myc, and Klf4. The authors designated these new cells induced pluripotent stem cells (iPS cells). Pluripotency is defined as a stem cell’s ability to develop into cells of any of the three germ layers. Embryonic stem (ES) cells are pluripotent cells that can self-perpetuate for long periods of time and are obtained by removing the inner cell mass from a blastocyst, which destroys the developing embryo. There has been controversy over using human ES cells in research, because of the ethical issues involved with destroying human embryos and the ambiguous beginnings of personhood. ES cells remain a popular research topic, however, due to their immense clinical promise. ES cells could be grown in cell culture, induced to differentiate into a certain tissue type, and then implanted into a patient with any number of diseases, including heart disease, Parkinson’s disease, spinal cord injury, and diabetes. However, if generic ES cells are used to grow these tissues, patients’ immune systems could reject the foreign tissue, which could be avoided by inducing the patients’ own cells back into the pluripotent stage and making healthy tissue from these cells.
ES cells and the cells in early embryos preserve their pluripotency by the action of transcription factors, such as Oct3/4, Sox2, and Nanog. The long-term self-perpetuation characteristic of ES cells is maintained by tumor-related genes including Stat3, E-Ras, c-Myc, Klf4, and β-catenin. Using this prior knowledge, Takahashi and Yamanaka chose 24 candidate genes to test for their ability to revert differentiated somatic cells into a pluripotent state. To detect pluripotency, they used a G418 resistance assay, in which they used homologous recombination to insert β-galactosidase and neomycin resistance genes fused in a βgeo cassette into the Fbx15 gene. When ES cells had the genotype Fbx15βgeo/βgeo, they could tolerate concentrations of G418 up to 12 mg/ml, while somatic cells with this same genotype were susceptible to much lower G418 concentrations of 0.3 mg/ml. Since Fbx15 is only actively transcribed in ES cells and early embryos, activation of Fbx15 after the addition of one or a combination of the 24 candidate genes would indicate that somatic cells had been induced to become pluripotent and could be detected by tolerance of normal amounts G418 in the medium. G418 is related to Gentamicin and inhibits eukaryotic translation (Invitrogen)
Results
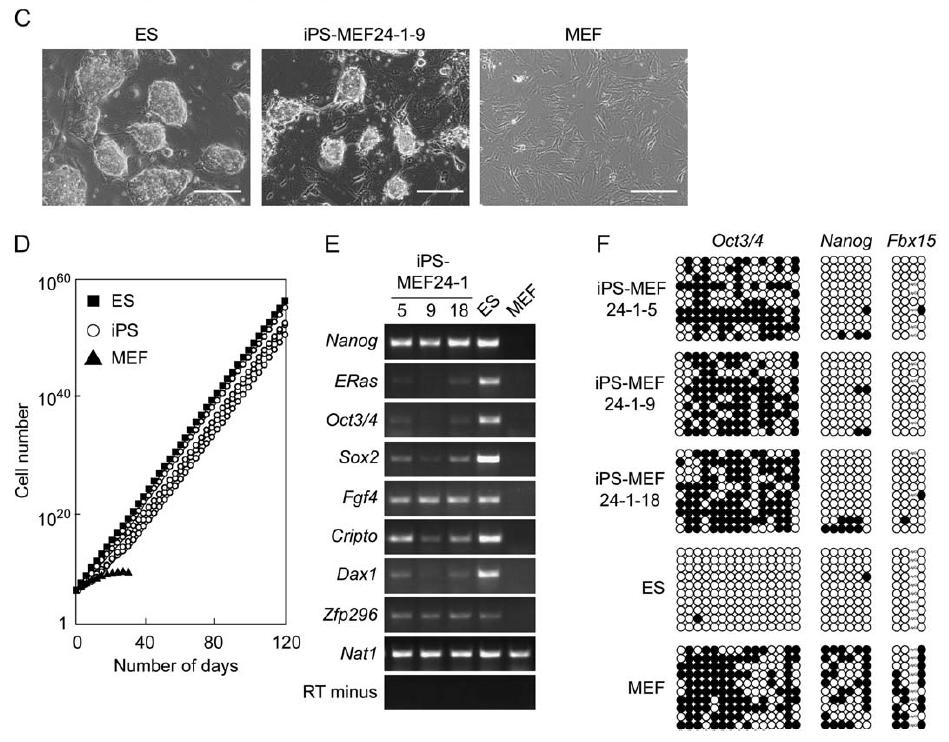
Figure 1A shows the retroviral transduction system that the researchers used to add the candidate genes one at a time into Fbx15βgeo/βgeo mouse embryonic fibroblasts (MEFs), which were grown on STO feeder cells in ES cell culture medium mixed with a normal concentration of G418 (0.3 mg/ml). Figure 1B demonstrates that no neomycin resistant colonies were observed when the candidate genes were added individually, which demonstrated that more than one gene is needed to turn on the Fbx15 promoter. In comparison, 22 G418-tolerant colonies were obtained when the 24 candidate genes were transduced into a fibroblast together, of which 5 had the circular shape, larger than normal nucleoli, and small amount of cytoplasm that physically characterize ES cells. When the experiment was duplicated, four colonies appeared to be iPS cells with the same morphology and prolonged proliferation as ES cells. These cells were called iPS-MEF24, which signified that they were induced pluripotent stem cells made from MEFs that were transduced with 24 genes (Figure 1C). Prolonged proliferation was demonstrated in Figure 1D through a growth curve, on which the lines for the iPS cell clones were indistinguishable from the line for ES cells and continued proliferating linearly for more than 120 days, but the MEF cells reached a horizontal asymptote and stopped proliferating before 40 days. Figure 1E shows that genes normally transcribed in ES cells, such as Oct3/4, Nanog, E-Ras, Cripto, Dax1, Zfp296, and Fgf4, were found by reverse transcription PCR (RT-PCR) to also be active in iPS-MEF24 cells. Bisulfite genomic sequencing of the iPS-MEF24 cells (Figure 1F) revealed demethylation in some promoters, including Nanog and Fbx15, which indicates that these genes were being expressed, while the Oct3/4 promoter was still methylated, which demonstrates reduced expression of this gene. Thus, only a mix of a few of the candidate genes is needed to induce pluripotency.
Figure 1. iPS-MEF24 clones are physically similar to ES cells, they have largely indistinguishable growth curves, they express several ES cell markers (including Nanog, Sox2, Fgf4, Cripto, Dax1, and Zfp296), and their Nanog and Fbx15 promoters are largely demethylated, which is similar to the ES methylation pattern, although their Oct3/4 promoters are still highly methylated.
Takahashi and Yamanaka removed one candidate gene at a time to distinguish which were essential to induce G418 resistance, and 10 of the 24 genes (designated 3, 4, 5, 11, 14, 15, 18, 20, 21, and 22) were shown to be critical due to lack of colony formation upon their removal (Figure 2A). As further evidence for these 10 genes being essential, there were more iPS colonies when MEFs were transduced with them than with all of the 24 combined (Figure 2B). One gene at a time was removed from this group, and genes 14 and 20 (Oct3/4 and Klf4) were found to be critical for colony formation. No colonies resistant to G418 formed upon the removal of these two factors. Few G418-tolerant colonies formed when gene 5 (Sox2) was taken out of the transduction mixture, and the colonies that formed when gene 22 (c-Myc) was removed no longer had morphologies similar to ES cells (Figure 2B). Thus, Oct3/4, Klf4, Sox2, and c-Myc are crucial to induce MEFs to become iPS cells. In fact, transducing MEFs with just these four genes formed four iPS lines (designated iPS-MEF4), while transduction with the 10 essential genes created five iPS-MEF10 colonies. Oct3/4, Klf4, Sox2, and c-Myc are, therefore, necessary and sufficient for iPS cell formation.
As shown in Figure 2C, the authors could not find two-gene transduction combinations that would produce colonies that could tolerate G418. Three-gene combinations (iPS-MEF3 cells) were only moderately more successful. Mixtures of Oct3/4, Sox2, and c-Myc and Klf4, Sox2, and c-Myc each formed one small G418-resistant colony that did not proliferate in future culture passages. A mixture of Oct3/4, Sox2, and Klf4 produced 36 colonies that tolerated G418 but that no longer physically resembled ES cells. A transduction mixture of Oct3/4, Klf4, and c-Myc yielded 54 G418-tolerant colonies, of which six proliferated in culture for several passages but had rough borders instead of the smooth, round appearance of ES cells and the iPS-MEF4 and iPS-MEF10 cells (Figure 2D).
To determine if iPS cells actively transcribed the genes for ES cell markers, Takahashi and Yamanaka ran RT-PCR with primers specific for the endogenous genes (Figure 3). The cell marker genes tested were Ecat1, Esg1, Nanog, ERas, Gdf3, Oct3/4, Sox2, Fgf4, Cripto, Dax1, Zpf296, Slc2a3, and Nat1. The iPS-MEF4 and iPS-MEF10 cell lines transcribed most of the markers, except Ecat1, and therefore, they closely resemble but are still distinguishable from ES cells. The majority of the iPS-MEF3 colonies failed to transcribe many of the marker genes, including Ecat1, Esg1, Oct3/4, and Sox2. Figure 4 demonstrates that gene expression analysis using DNA microarrays revealed Myb, Kit, Gdf3, and Zic3 were upregulated in both ES and iPS cells. iPS-MEF3 cells were once again distinguishable from ES, iPS-MEF4, and iPS-MEF10 cells, as they did not efficiently upregulate Dppa3, Dppa4, Dppa5, Nanog, Sox2, Esrrb, and Rex1, which could explain why these cells were not pluripotent. As further evidence that iPS cells are not identical to ES cells, only ES cells upregulated Dnmt3a, Dnmt3b, Dnmt3l, Utf1, Tcl1, and the gene for the LIF receptor.
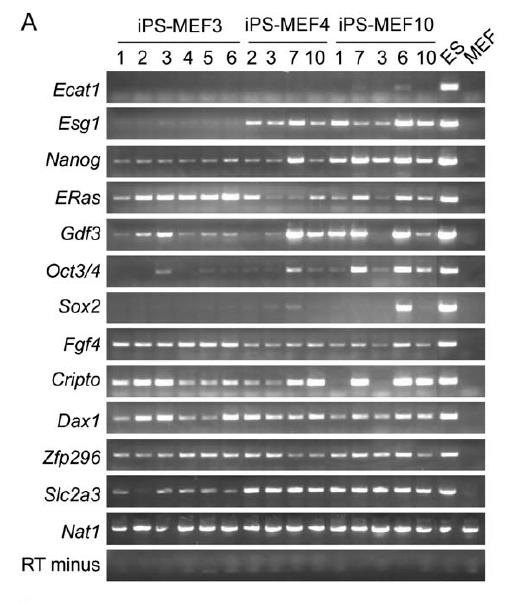
Figure 3A. The iPS-MEF4 and iPS-MEF10 cell lines transcribed most of the ES cell markers, except Ecat1, and therefore, they closely resemble but are still distinguishable from ES cells. The majority of the iPS-MEF3 colonies failed to transcribe many of the marker genes, including Ecat1, Esg1, Oct3/4, and Sox2.
As shown in Figure 5, the authors subcutaneously injected nude mice with the different iPS cells in order to test their pluripotency by tumor formation. Tumors formed with five of the iPS-MEF10 colonies, three of the iPS-MEF4 colonies, and six of the iPS-MEF3 colonies. Of these tumors, iPS-MEF10 colonies number 3 and 6 and iPS-MEF4 colonies 2 and 7 were truly pluripotent, as they had cells from all three germ layers, containing nervous tissue, connective tissue, and epithelium (Figure 5A). All iPS-MEF3 tumors consisted of undifferentiated cells and, thus, these cells were not pluripotent (Figure 5C). Figure 5B confirms the pluripotency of iPS-MEF4-7 through immunostaining the tissues from the teratoma for smooth muscle actin and neural-specific proteins GFAP and βIII tubulin. Figure 5D demonstrates that when iPS-MEF4-7 and iPS-MEF10-6 were stimulated to differentiate in vitro, they formed cells from all three germ layers. Figure 5D is immunostaining, in which primary antibodies for smooth muscle actin, α-fetoprotein, and βIII tubulin were used and then the secondary antibodies were made red by Cy3. For the α-fetoprotein in iPS-MEF10-6, a green Alexa 488 secondary antibody was used instead of the red Cy3 secondary antibody.
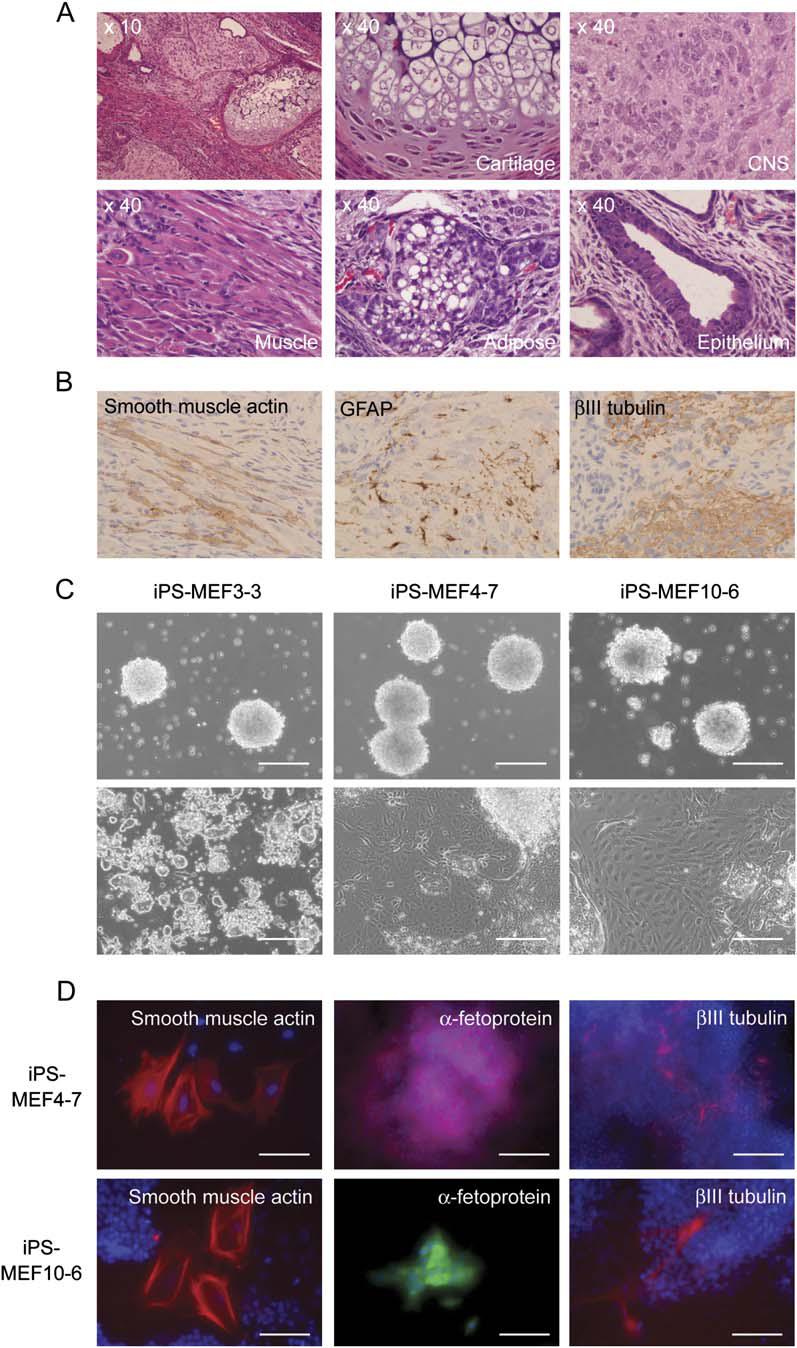
Figure 5. iPS-MEF10 clones number 3 and 6 and iPS-MEF4 clones 2 and 7 were truly pluripotent, as they had cells from all three germ layers, containing nervous tissue, connective tissue, and epithelium (confirmed by microscope images in (A), immunostaining of the teratomas in (B), and immunostaining of cells differentiated in vitro in (C)). All iPS-MEF3 tumors consisted of undifferentiated cells and, thus, these cells were not pluripotent.
As illustrated in Figure 6, to further demonstrate that Oct3/4, Klf4, Sox2, and c-Myc are necessary and sufficient to produce iPS cells, the researchers transduced them into tail-tip fibroblasts (TTFs) from four 7-week-old male mice with the genotype Fbx15βgeo/βgeo and one 12-week-old female Fbx15βgeo/βgeo mouse that also produced GFP from the CAG promoter. Three iPS cell clones (iPS-TTF4) were made from the male mice, and six clones (iPS-TTFgfp4) were established from the female mouse. These clones were all physically identical to ES cells (Figure 6A), and iPS-TTFgfp4 clones 3 and 7 transcribed all ES marker genes, some at higher levels than others (as demonstrated by RT-PCR in Figure 6B). Two of the iPS-TTF4 colonies and six of the iPS-TTFgfp4 clones were injected into mice and were all pluripotent, forming tumors with cells from all three germ layers (Figure 6D). When iPS-TTFgfp4-7 and iPS-TTFgfp4-3 were injected into blastocysts, two of the 18 embryos obtained from iPS-TTFgfp4-3 and three of the 22 embryos obtained with iPS-TTFgfp4-7 were positive for GFP (Figure 6C). 27 mice were born from these two microinjections, none of which were chimeras. Thus, the GFP+ iPS cells did not differentiate into germ cells.
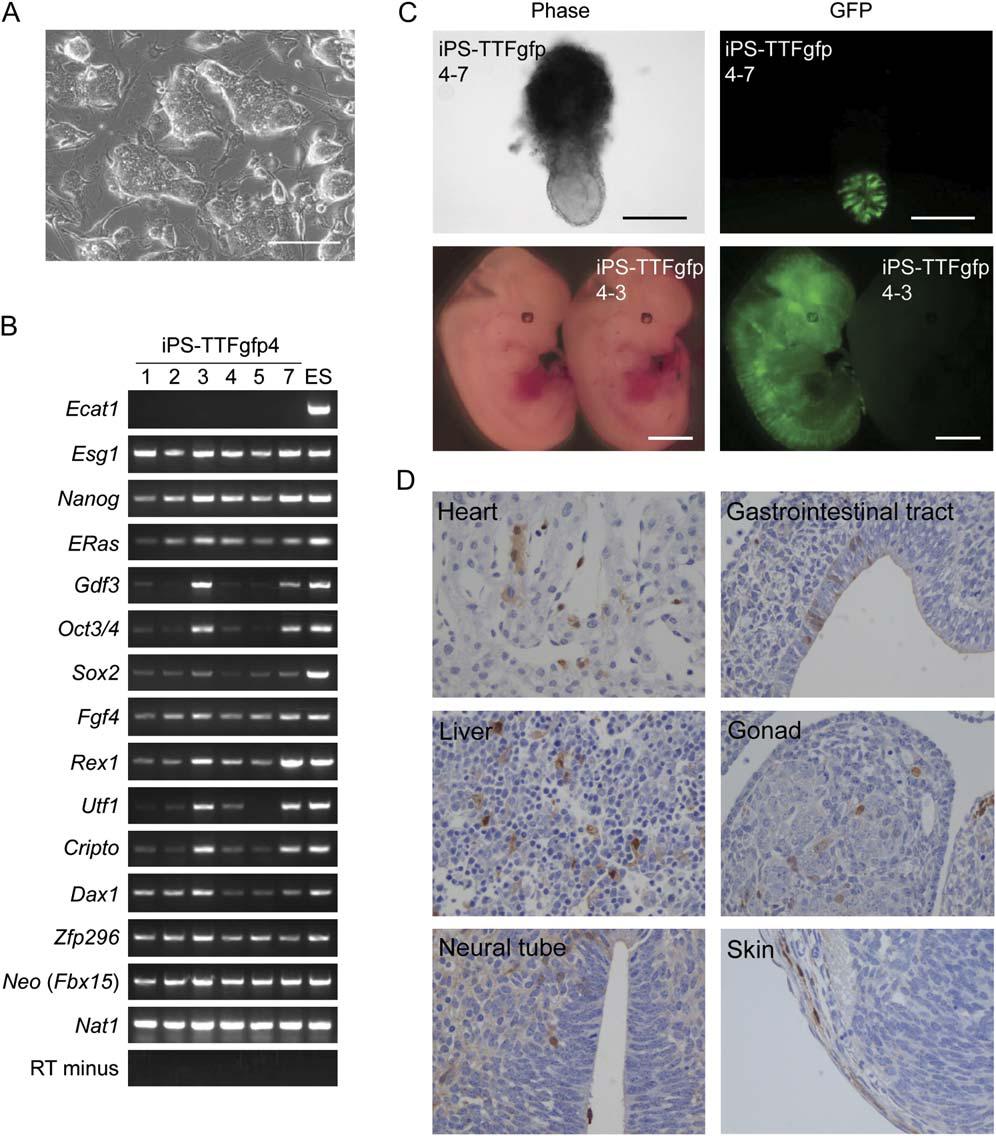
Figure 6. A: iPS-TTFgfp4 clones were all physically identical to ES cells. B: RT-PCR showed that iPS-TTFgfp4 clones 3 and 7 transcribed all ES marker genes, some at higher levels than others. C: When iPS-TTFgfp4-7 and iPS-TTFgfp4-3 were injected into blastocysts, two of the 18 embryos obtained from iPS-TTFgfp4-3 and three of the 22 embryos obtained with iPS-TTFgfp4-7 were positive for GFP. D: Two of the iPS-TTF4 colonies and six of the iPS-TTFgfp4 clones were injected into mice and were all pluripotent, forming tumors with cells from all three germ layers.
Figure 7 is mostly a summary of all of the other figures, this time comparing the iPS-TTFgfp4 clones to the iPS-MEF4 and iPS-MEF10 clones and ES cells. Figure 7A shows a Western blot of protein expression in iPS-MEF4-7, iPS-MEF10-6, iPS-TTFgfp4-3, iPS-TTFgfp4-7, ES, and MEF cells. β actin is a loading control, and the same level of protein is loaded in every lane. Three out of the four iPS proteins (Oct3/4, Sox2, and Klf4) are expressed about the same amount in all the iPS clones as in ES cells. c-Myc is expressed at about the same level in all the iPS clones and ES cells and at a slightly lower level in MEF cells.Nanog is only translated to a substantial degree in iPS-MEF10-6 and ES cells, which makes sense due to the fact that it was one of the 10 factors added to iPS-MEF10-6 and was not added to the iPS-MEF4 or iPS-TTFgfp4 clones. ERas is translated more in iPS-MEF10-6 than in ES cells, but it is also translated to a lesser degree in all the other iPS clones. p53 is translated more in MEF cells than in ES cells or in any of the iPS clones, although it is translated to some degree in all of them. Figure 7B shows that the RNA and protein for Oct3/4, Sox2, and Nanog are always expressed more in undifferentiated iPS and ES cells than in differentiated iPS and ES cells and are not expressed at all in MEF or TTF cells. Figure 7C is a Southern blot that demonstrates about 20 transgenes from the retrovirus were integrated into the DNA of each iPS clone. Figure 7D is a visually normal karyotype developed from a developing embryo that had been microinjected with iPS-TTFgfp4.
Critical Analysis
The results in this paper clearly support the authors’ claims. The figures are very expansive and generally have four or more parts to them. Their RT-PCR images always have a clear positive control (Nat1) and an RT minus negative control. Also, every RT-PCR image contains a lane for ES cells and MEFs to see clearly which genes are normally actively transcribed in these cells. The ES lane allows for comparison of the normal expression level of the different marker genes to that in the iPS cells. The microscope images are clear, and the iPS cells obviously look morphologically much more similar to ES cells than to MEF cells. The authors always support their microscope pictures with other experimental evidence, such as the RT-PCR gel images, bisulfite genomic sequencing images to determine promoter methylation status and growth curves.
Furthermore, the authors do not overstate their findings. They mention several times that while they are similar, iPS cells are not identical to ES cells. The authors do not rule out the possibility that they neglected additional necessary genes for ES cell maintenance, such as Ecat1. They admit that one problem in their research was that a very small proportion of fibroblast cells became iPS cells. The authors give a few explanations for this observation. This issue could be due to inefficiencies with the retrovirus expression system. Or more disturbingly, rare tissue-specific stem cells in the fibroblast cultures could have become iPS cells when they took up the four ES genes. If this were the case, the authors’ argument is weakened, because it is not fully differentiated cells reverting to a pluripotent state but multipotent stem cells becoming pluripotent stem cells. The authors tested this possibility by adding Oct3/4, Sox2, Klf4, and c-Myc to bone marrow cells, which have a greater proportion of multipotent stem cells than do skin cells, and iPS cells formed at a similarly low efficiency to the fibroblast cultures. Thus, tissue stem cells probably are not the source for iPS cells. Another possible explanation for the low frequency of iPS cell formation is that the four necessary genes must be expressed at certain, very narrowly defined levels, and only a few cells transcribe the genes in exactly the right amounts. Prior knowledge supports this explanation, because ES cells will differentiate if the cellular Oct3/4 protein concentration alters by as little as 50%. Finally, the four genes may induce chromosomal changes, such as alterations in gene silencing, that must occur in culture in order for iPS cells to form.
Potential Next Experiments
An obvious next experiment to follow this work would be to use the retrovirus expression system in human somatic cell cultures to add Oct3/4, Sox2, Klf4, and c-Myc and see if iPS cells form. A knockin construct such as the Fbx15-βgeo system used here could be applied in this human cell culture, or a GFP gene could be added under the control of a promoter only active in ES cells, such as that for Ecat1 or Esg1, so iPS cells would fluoresce green in culture. Then RT-PCR with endogenous protein-specific primers could confirm the expression of ES cell genes in these iPS cells.
Another next step would be to find another gene besides c-Myc that would maintain the proliferation potential of iPS cells, because c-Myc is not clinically applicable. This protein has many downstream target genes, including those for histone acetyltransferase (HAT) complexes, which are necessary for overall chromosomal structure. The human genome may have as many as 25,000 sites for c-Myc binding, so a less globally-active protein may be preferable for the clinical use of iPS cells. Other tumor-related genes, such as Stat3, E-Ras, or β-catenin could be transduced with the other three genes to see if iPS cells formed with the same relative efficiency. For initial experimental purposes, the same Fbx15 G418 resistance assay explained here could be used to evaluate the effectiveness of these other genes.
A final future experiment would be to determine whether iPS cells are preserved by the transgenes transcribed from the retrovirus that was transduced into the mouse cells. Takahashi and Yamanaka found that the endogenous expression of Oct3/4 and Sox2 proteins was very low in iPS cells. Obviously, it may not be clinically practical to constantly have a retrovirus in cell culture in order to produce enough iPS cells to differentiate into a transplantable tissue or organ. To test this question, the genes for the four factors in the retrovirus could be put under control of an inducible expression system, such as the tetracycline inducible expression system Flp-In T-REx™ (Senkel et al., 2009). In this system, tetracycline would initially be present in the medium to allow the genes from the retrovirus to be transcribed and translated to induce the formation of iPS cells from somatic cells. Then tetracycline could be removed between passages to see if the iPS cells could be maintained by their endogenous genes or if they differentiated. Based on the preliminary results obtained by Takahashi and Yamanaka, the iPS cells could be expected to differentiate once the transgene expression was removed.
Works Cited
G418, Geneticin. [Internet]. 2010 [cited 2010 Apr 28]. Invitrogen. Available from: http://www.invitrogen.com/site/us/en/home/Products-and-Services/Applications/Cell-Culture/Transfection/Selection/g418.html?cid=covinvggl89200000000097s&
Senkel S, Waldner C, Ryffel GU, Thomas H. 2009. Improved conditional expression systems resulting in physiological level of HNF4α expression confirm HNF4α induced apoptosis in the pancreatic β-cell line INS-1. BMC Research Notes 2:210.
Takahashi K, Yamanaka S. 2006. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126: 663-676.