This web page was produced as an assignment for an undergraduate course at Davidson College.
My Favorite Yeast Proteins
MCA1p and YOR205Cp
Introduction
On previous websites, information about the genomic information and the expression data concerning MCA1 and YOR205C. These data showed the availability of public information about yeast genes that can determine the direction of new resource into yeast gene functions. On these previous websites, data for an unannotated yeast gene (YOR205C) were presented in order to hypothesize about the biological and molecular function of this molecule. On this site, this information will be combined with information from proteomics databases that analyze protein-protein interactions and the properties of YOR205Cp and MCA1p to further determine how these proteins operate within a yeast cell. But the data gathered from these databases only allows hypotheses to be formulated about the function of non-annotated yeast genes. These hypotheses must then be tested at the bench to validate or refute them. Therefore, after presenting a final hypothesis about the function of YOR205Cp within yeast cells, I will propose directions for further research and experimentation into this gene that will provide real data about the gene's function.
MCA1p - Proteomics Information
Previous analyses of MCA1 can be found here and here. Briefly, gene and expression data showed that MCA1 was a caspase involved in the yeast apoptotic pathway, which controls and signals programmed cell death in response to a variety of signals including harsh environmental stress and DNA damage. The GO terms associated with this gene include localization to the nucleus, caspase activity, and apoptosis. Expression data showed that the protein was expressed alongside telomere maintenance genes, other genes in the apoptotic pathway, and environmental stress response factors.
Basic Protein Information
The NCBI protein information about MCA1 showed that the MW of MCA1 is 47851 Da (NCBI). The SGD database showed that the protein's Isoelectric point, or pI, is 5.09, but lists the MW as 47982 (SGD), which is relatively close to the NCBI value. In addition to this general information, however, the SGD database provides information about the physical interactions of MCA1p.
2D Gel Localization
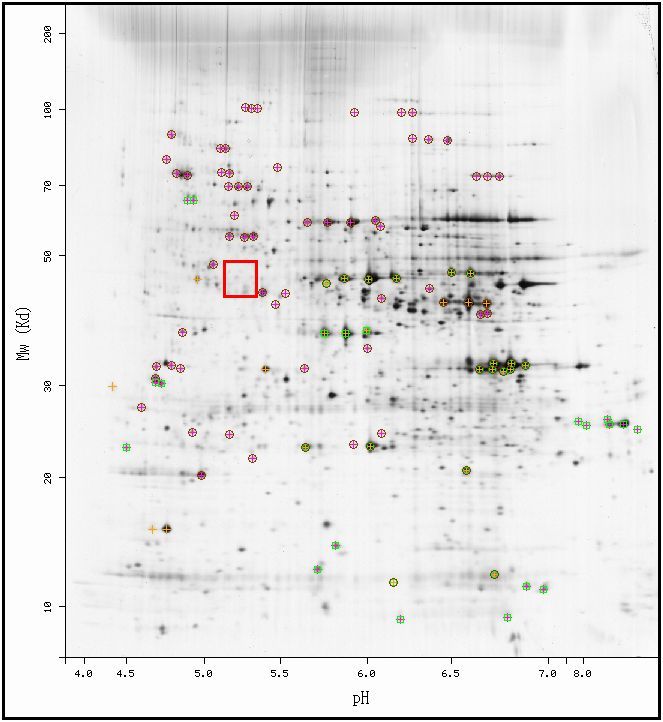
From the MW and pI, one can determine the position of the protein on a 2D-gel found in the ExPASy database. The picture below shows a 2D gel electrophoresis diagram of the yeast genome. Each spot on the diagram corresponds to the existence of protein in the gel, since each protein can be separated by pI (on the horizontal axis) and by MW (on the vertical axis). According to the data above, the MCA1 protein should exist somewhere inside the red box.
|
| Figure 1. 2-D Gel Electrophoresis diagram of all identified proteins in the yeast genome. Permission pending from ExPASy. |
| As seen in the above figure, the space in the red box, corresponding to the predicted area for MCA1, contains some dark marks (signifying proteins) but does not include any idenfified spots, or spots with crosses or circles around them. Therefore, this database does not further the hypotheses presented about MCA1 from the data presented above. |
Protein Interactions - Yeast Two-Hybrid and Affinity Capture MS Data
Two related methods provide high-throughput methods of determining the interactions of proteins in a genome. Both of these methods - Yeast Two-Hybrid and Affinity Capture MS - rely on detection of protein complexing in laboratory-grown yeast cells in which one protein is used as "bait" to draw out reactions with numerous "prey" proteins. In Yeast Two-Hybrid, or Y2H, bait proteins are attached to one half of a transcription factor for a reporter gene, such as GFP. These proteins are inserted into cells along with multiple types of prey proteins that are complexed with the other half of the TF. When the bait complexes with a prey protein, the halves of the transcription factor are connected and the reporter gene is produced in those cells (Campbell and Heyer, 2006). More information on Y2H, including an animation, can be found here. The Affinity Capture/MS method works in a similar way in that it detects protein interactions, but it does not use TFs to detect protein complexing. In this method, protein interactions are determined by adding proteins to yeast cells, allowing these proteins to complex (if applicable), then isolating them using antibodies and analyzing the complex using Mass Spectrometry (SGD). (More on this method at the SGD glossary). This method has the advantage of not requiring the addition of a protein segment onto each of the bait and prey proteins, which could disrupt potential complexes. Both of these methods are used in yeast cells to determine interactions between different yeast proteins, but each contains flaws. Because proteins are added to yeast and not studied in vivo, some protein-protein interactions may not necessarily occur normally in a yeast cell. For example, two proteins could interact in a Y2H experiment but could be active during different parts of the cell cycle and would therefore never interact in an actual cell. Despite this lack of certainty, these complexing experiments provide good starting points for determining protein interactions and are some of the most well-used, high-throughput methods of studying yeast proteins.
SGD shows that MCA1 interacted with one protein in an Affinity Capture/MS experiment and eight more in Y2H experiments, as seen in the following table:
Protein Name |
Interaction Type |
Summary of Function (from gene pages) |
Affinity Capture/MS |
tRNA binding protein that directs the placement of tRNA |
|
Y2H |
Protein required for “nucleotide excision repair” (a DNA repair enzyme) |
|
Y2H |
Mediator/regulator of RNA Polymerase II activity (mediates transcription) |
|
Y2H |
Cell wall biosynthesis protein |
|
Y2H |
Protein kinase involved in cell signaling that controls growth rate as a function of glucose availability |
|
Y2H |
Binds to mRNA and interacts with mRNAs associated with the cellular membrane |
|
Y2H |
Protein associated with the membrane of vesicles in the Golgi (exocytosis…) |
|
Y2H |
“Nucleotide exchange factor,” but it’s a pseudogene |
|
Y2H |
Overproduction of this protein causes production of the PIN prion |
Table 1. Proteins interacting with MCA1, either as bait or prey in Y2H and AC/MS experiments. Functional information summarized from the links to each gene. Table adapted from <http://db.yeastgenome.org/cgi-bin/phenotype/phenotype.pl?dbid=S000005723&type=allinteractions#physical>.
| These proteins in the table constitute proteins that interacted with MCA1 in a laboratory setting of yeast cells; interactions could be indicative of protein-protein interactions but could also constitute artifacts of the experiment. Some of these proteins could be predicted to interact with MCA1. These include proteins that are involved in DNA repair and transcription, since an abundance of DNA errors can cause the initiation of apoptosis (and this interaction provides the initial signaling interaction). Others could be implicated because of a possible role in initiating apoptosis due to lack of resources, such as YAK1. This adaptation could cause cells without enough resources to undergo apoptosis, thereby saving those resources for other yeast cells in the colony who could grow on those limited nutrients. However, there are proteins included on this list that can be considered artifacts of the experimental tests. For example, it seems very unlikely that a pseudogene (SDC25) would interact with MCA1, since it does not produce a functional enzyme in S. cerevisiae. This finding calls into question the other Y2H interactions listed below – although some of the interactions ‘make sense,’ do these proteins actually interact in the cell? It is impossible to definitively tell from the data above, but it certainly provides an indication toward possible interactions of MCA1 within the cell. Finally, the list above is probably not exhaustive; chances are other proteins interact with MCA1 to activate it or signal it to begin the apoptotic cascade. |
Other online databases provide further Y2H study results. The following table was derived from the Yeast Resource Center and compiles information on additional Y2H interactions and the functions of the interacting proteins. In addition to this data, the Yeast Resource Center reported the same results listed in the table above, providing additional redundant evidence that these proteins interact with MCA1.
Protein Name |
Summary of Function (from SGD gene pages) |
Uncharacterized ORF |
|
Protein involved in the formation of the cytoskeleton |
|
Helicase and telomere maintenance protein |
|
Chromosome Segregation during Mitosis |
Much like the previous data, this one provides genes that would seem to interact with an initiator caspase. Genes that are involved in telomere maintenance and chromosome segregation, if errors are detected in these processes, could signal the cell to initiate apoptosis if the errors in these essential cellular processes are great. In addition, these data provide an additional method of determining the function of genes. An uncharacterized yeast ORF was detected as a prey protein when MCA1 was used as bait, which means that these two proteins could possibly interact. This would allow researchers to propose a function for YKR096w on the basis of its interactions with other proteins. |
Interactomes from the Database of Interacting Proteins
One database that provides information about protein-protein information in the form of interactomes, or graphs of interactions displayed in a node-and-edge format, is the Database of Interacting Proteins, or DIP. These interactomes show a graphical display of yeast gene interactions, thereby allowing one to view not only the interactions of a single protein but those of a network, creating linkages between proteins that interact with a common enzyme and functional linkages with whole processes inside the yeast cell. I searched the database for MCA1 and determined the following information about protein interactions from this site:
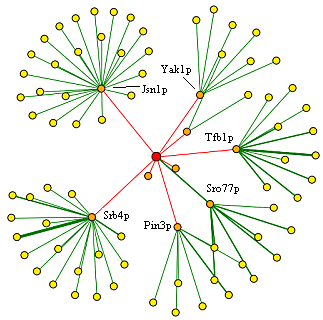 |
 |
| Figure 2. Protein interaction data for MCA1 from DIP. See legend in Figure 3 for details. Permission Pending from DIP. | Figure 3. Legend used to interpret above figure from DIP. Permission Pending from DIP. |
| These data show that there are a large number of proteins connected in some way to MCA1, either through primary interactions or secondary interactions. The proteins in orange, or the ones that represent first-shell nodes, consist of proteins listed in the tables above, from SGD and the Yeast Resource Center. The DIP database probably derives information from the same studies as both of these databases, therefore, since the same data is presented. This figure also shows a level of depth not seen in the other databases, however, because of the existence of secondary interactions in the interactome. The proteins with the highest degree (number of edges) are the ones that are involved in mRNA binding and transcription, such as Jsn1p. These enzymes could be transcription factors that bind to multiple other enzymes to function, or they could be components in a widely used process that is modulated by many other proteins. Overall, the data above show the large amount of primary and secondary interactions of MCA1p, as well as the extensive amount of linkage between different biological processes in the yeast cell. |
Other Database Searches
MCA1 not found in the PROWL database, the Protein Data Bank, or in the ExPASy 2D search database.
Interactomes displaying genes involved in yeast aging, degradation, and cellular membrane structure did not include the search term "MCA1."
Nothing found in the Fields Lab dataset of interactions or the TRIPLES database when searching using the terms "MCA1" or "YOR197w."
An entry for MCA1 was found in the KEGG database; however, this information had already been analyzed previously.
The YOR197w (MCA1) gene was not found in the PROWL database.
Conclusions
MCA1 protein interaction data presented data that expanded on gene sequence analysis and gene expression data. Y2H protein-protein interactions provided evidence that MCA1 complexes with a diverse range of proteins, including those involved in DNA damage repair, cell wall biosynthesis, telomere maintenance, and cell signaling. Many of these interactions would be expected from a protein that serves as the beginning of the apoptotic pathway in the cell; for instance, MCA1 could interact with enzymes in cases of extreme telomere shortening, high amounts of DNA damage, or cell wall damage and respond by being modulated by these proteins, beginning the process of apoptosis in the yeast cell. However, pseudogenes were also shown to hybridize with MCA1, providing evidence that some of these interactions may be artifacts and never actually occur in these organisms. It is possible that the pseudogene listed above is mislabeled and actually performs a cellular function, if it interacts with MCA1. However, it is more likely due to the nature of Y2H that the two proteins do not complex in vivo. Because of this uncertainty, high-throughput methods of testing hypotheses, such as Y2H, should be presented with the knowledge of this uncertainty. If possible, further experimentation should be performed on this protein to determine whether these interactions actually occur in vivo, or whether these interactions are simply artifacts of the Y2H or AC/MS experiment. Studies to test whether these proteins, such as MCA1 and JSN1, actually do interact in vivo is to detect whether they occur in the same places in the yeast cell and at the same points in the cell cycle. If they are active at different times or in different places in the cell, then chances are the results above for that pair of proteins are experimental errors. Without this information, one can hypothesized that many (though not all) of the genes listed in Tables 1 and 2 complex with MCA1 to change each molecule and perform some function within the yeast cell.
YOR205C Proteomics Information
Although these databases are useful for reiterating and comparing information for known genes, they can also be used in a predictive capacity. Previous websites (here and here) have used gene and expression databases to characterize a non-annotated gene, YOR205C, about which a relative lack of information is known. Up until this point, I have made the following hypotheses about YOR205C:
The only GO term available for the gene localized the protein to the mitochondria; however, this information is not enough to conclusively state that the gene is located there. Proteomics databases will help to further support or refute this hypothesis.
Sequence comparison and expression comparison show that the gene probably has something to do with RNA binding, possibly implicating it as an RNA Helicase. Since proteomics databases only display protein-protein interactions, a complex with RNA will not be verified on this site. However, if interactions of the protein link it to other genes involved in RNA binding or modulation of enzymes associated with RNA, these links would further this hypothesis.
Expression alongside many ribosome biogenesis molecules seems to indicate that YOR205C could be implicated in the formation of ribosomes. If the gene complexes with pieces of a ribosome, then it is highly likely that the gene modulates or interacts with the ribosome during formation, either by changing its shape or adding and connecting pieces to the ribosome.
To test these hypotheses, I will search for the protein in many proteomics databases, then propose experiments to test these hypotheses.
Basic Information
The SGD Database provides some basic information about the YOR205C gene:
 |
| Table 3. General protein information about YOR205C. Permission pending from SGD. |
These data show that YOR205C is larger than MCA1 and falls on the higher end of the spectrum of isoelectric points for proteins. From this information, one can search for the gene on the ExPASy 2D gel image to determine if additional information can be characterized from its position on the gel.
2D Gel Localization
The 2D Gel from ExPASy provided the following information:
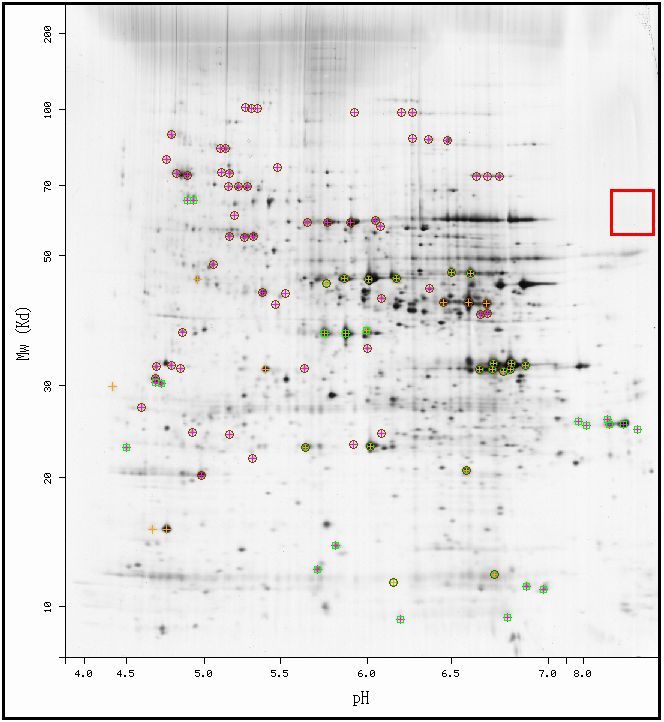 |
| Figure 4. Figure 1. 2-D Gel Electrophoresis diagram of all identified proteins in the yeast genome. Permission pending from ExPASy. |
| Although the protein was expected to be in the red box shown above, there seem to be no proteins localized in the red box, evidenced by the lack of dark signals on the 2D image. Although this could be due simply to the poor resolution of the image, it is possible that this image does not show the protein. The image claims to display all "identified proteins" in the yeast genome (ExPASy, 2006), the lack of characterization of YOR205C could have caused it to be left out of the analysis. |
Protein Interactions - Yeast Two-Hybrid and Affinity Capture MS Data
SGD showed only one interaction between YOR205C and other proteins in the yeast genome. The yeast protein MRPS5p interacted in an affinity capture/MS reaction with FMP38p, an alternative name for YOR205C (SGD YOR205C gene, 2006). Upon further investigation, MRPS5 was determined to be a gene that codes for a section of the mitochondrial ribosomal protein, as seen in the data below. MRPS5 was also described as a "Mitochondrial ribosomal protein of the small subunit" from a paper in the Journal of Biological Chemistry (Saveanu et al., 2001).
 |
| Table 4. Description of GO terms for MRPS5, from SGD. Permission Pending from SGD |
This protein interaction provides evidence for multiple conclusions about YOR205C. Previous pages have suggested from the GO terms associated with the gene that the protein product is located in the mitochondria; interactions with only a mitochondrial protein show that this gene indeed could produce a mitochondrial protein, if this interaction is verified. In addition, the protein that supposedly interacts with YOR205C is a RNA-binding protein, indicating that YOR205Cp could also interact with nucleic acids or with processes involving RNA, such as translation (like MRPS5). If this interaction is correct, it would implicate this gene in a translational control mechanism or a protein that modifies or interacts with the process of translation. Alternatively, however, this interaction with a ribosomal subunit could further the hypothesis that the gene interacts with segments of the ribosome during ribosome biogenesis. By modifying MRPS5, the YOR205C protein could play its role in the formation of the mitochondrial ribosomes. However, this protein-protein interaction could also be an artifact. If YOR205C is not produced in the mitochondria after all, this protein-protein interaction would never occur outside a laboratory setting in vivo. If the interaction is indeed an artifact, then the conclusions made above are less substantial. |
Interactomes from the Database of Interacting Proteins
A search in the Database of Interacting Proteins for YOR205C revealed the following interactomes, which show the gene (symbolized by its alternative name FMP38) only having an interaction with one other protein, thus reiterating the data above. I found the following information about YOR205C interactions from this page:
 |
|
| Figure 5. Protein interaction data for YOR205C from DIP. See legend in Figure 6 for details. Permission Pending from DIP. | Figure 6. Legend used to interpret above figure from DIP. Permission Pending from DIP. |
As seen on this map, YOR205C has only one known interactions in the database, which links it to three other proteins in secondary interactions. This lack of interactions is not surprising since this gene has not been well-studied and is probably an indication of lack of research rather than lack of interactions. The one protein that YOR205C is linked to was MRPS5p, the same protein analyzed above. |
The proteins that are linked to MRPS5p in the above database are listed below, with their functions derived from SGD:
Protein Name |
Summary of Function (from SGD Gene Pages) |
Small subunit protein of the mitochondrial ribosome; interacts with COX3 |
|
Small subunit protein of the mitochondrial ribosome |
|
tRNA-binding segment of the mitochondrial ribosome |
The fact that all three proteins listed above constitute components of the mitochondrial ribosome is expected, since MRPS5 produces a segment of the same macromolecule that is responsible for translation. What is interesting, however, is the exclusion of other proteins that interact with the ribosome. For instance, if MRP1 interacts with COX3, why is this protein not on the interactome? However, the graph shown above only goes so far away from YOR205C for the sake of simplicity and to prevent drawing conclusions about the relatedness of distantly-related proteins that may not interact directly. Therefore, information in this database indicates that YOR205Cp interacts with the mitochondrial ribosome, which, if true, would signal that this protein is in fact localized to the mitochondrial and is involved in modifying or interacting with translation. |
TRIPLES Protein Data
The TRIPLES website provides information on gene promotion and production in yeast cells. By inserting transposons with the LacZ gene encoded, researchers were able to insert reporter genes in various places throughout the yeast genome, including places downstream of real promoters (Campbell and Heyer, 2006). In some cells, transposons were inserted into YOR205C, disrupting that gene but allowing LacZ to be produced by the YOR205C promoter. The following figure shows that under conditions of both vegetative (normal) growth and under sporulation, YOR205Cp is produced heavily:
 |
| Figure 7. TRIPLES data for YOR205C. Permission pending from TRIPLES (Kumar et al., 2000). |
These data show that this gene is expressed during both normal conditions and conditions of rapid change in the cell. Although many genes are expressed during all times within a yeast gene, it makes sense that a gene regulating ribosome biosynthesis would be active at all times, since new ribosomes to facilitate translation are probably needed during all stages of yeast growth and life. Therefore, these data support the conclusion that YOR205C is involved in ribosome synthesis, but do not necessarily rule out any other functions for the gene besides highly specialized ones, such as specific sporulation responses.
Other Database Searches
A search of PDB returned no results for either “FMP38” or “YOR205C.”
The YOR205C gene was not found in the PROWL database.
Nothing found in the Fields Lab dataset of interactions.
The KEGG database included an entry for YOR205C but provided no new information.
Interactomes displaying genes involved in yeast aging, degradation, and cellular membrane structure did not include the search terms “FMP38” or “YOR205C.”
Conclusions
Overall, the yeast protein information presented above supports the conclusions made on previous websites (accessed here). Most of the hypotheses presented above - specifically, targeting YOR205C to the mitochondria and interactions with ribosomes - received further evidence, though proteomics information did not provide data showing that YOR205C is an RNA helicase. The Y2H data is presented in two separate fashions - as raw data in the SGD database and in graphical format in the DIP. Both databases show that YOR205C complexes with MRPS5p, a piece of the mitochondrial ribosome. Although the databases present the same data, it is probable that they compiled data from the same source; therefore, each database cannot be said to add additional evidence, only show the evidence in multiple fashions. In addition to the basic information, the DIP provides information about secondary interactions, or proteins that interact with proteins that complexed with YOR205Cp in Y2H experiments. These secondary interactions include other pieces of the mitochondrial ribosome, further supporting the association of the non-annotated gene with mitochondrial translation or ribosome biogenesis. Unfortunately, many of the proteomics databases did not include information for this gene, thereby only providing me with a small dataset on which to further the hypotheses made in previous websites.
When combined with previous data, the findings presented above provide enough evidence to hypothesize that YOR205Cp is indeed localized to the mitochondria, since it presumably complexes with mitochondrial ribosomal subunits. This complex along with expression data also further the hypothesis that the gene is a player in ribosome biogenesis. Although no data was presented on this site to show that the gene codes for an RNA helicase, the links to translation (which involves RNA) and formation of translational enzymes, as well as the sequence analysis data, point to the possiblity that the gene could include an RNA binding site. From the data presented in the preceding databases, both here and in previous websites, I hypothesize that YOR205C codes for a mitochondrial enzyme with a possible RNA binding site that plays a role in ribosomal subunit processing and mitochondrial ribosome biogenesis.
Experiments
Of course, the hypothesis presented above is just that - a hypothesis. To test this proposed function, experiments must be performed on this protein to determine its function, location, and process in the yeast cell. Experiments can be designed to test all parts of the hypothesis presented in the conclusions.
First, the location of the protein can be tested through a variety of methods, the easiest of which could be in situ hybridization. In this experiment, one would isolate yeast cells and inject them with antibodies that would bind to YOR205Cp. These antibodies would be bound to a secondary flourescent antibody that could then be detected. By detecting where in the cell the flourescent protein binds, one could easily determine the location of YOR205C in vivo. However, this method presents problems both in antibody production and targeting to the mitochondria. To make these antibodies, one could expose a pure form of the protein to mice or rabbits to create an immune response, then isolate the antibodies produced by the host animal to use for analysis. Unfortunately, though, the "pure" form of the protein could be structurally different from the protein in yeast cells, due to post-translational modifying and complexing with other molecules lost in the purification process. In this case, rabbit antibodies would not bind to the active form of the protein in the yeast cell. Additional problems occur when trying to inject antibodies into yeast cells - would they be able to pass through the mitochondrial membrane? Therefore, it would be beneficial, if possible, to isolate the mitochondria from yeast cells and perform hybridization on just this component. In this case, localization to the mitochondria would be signaled by any flourescence at all, as opposed to flourescence in a single part of the cell.
Because of the problems associated with in situ hybridization in this case, it might be possible to detect the location of the molecule another way. If one could tag the protein MRPS35p with a labeled antibody or other detection device, one could inject cells with this protein to determine where the non-annotated gene is located. It is already established that MRPS35p complexes with YOR205Cp inside a yeast cell. In addition, MRPS35p is already targeted to the mitochondria and could therefore get through those membranes. Through this method, one could detect the location of YOR205Cp using an altered hybridization technique that employs labeled mitochondrial ribosome subunits instead of antibodies that might not be specific for the active form of my favorite yeast protein.
Additional experiments would be required to test the other hypotheses presented above. A slightly modified version of yeast two-hybrid experiments could be used to further determine the role of YOR205C with ribosomes. The ribosomal subunit in the mitochondria is made up of many subunits; each subunit could be attached to one part of a transcription factor (or to multiple transcription factors) such that when all the parts of the ribosome come together correctly, the transcription factors line up and a reporter gene is produced by the yeast cell. Although transcription factors might only split into two pieces, a reporter promoter could be used such that additional TFs are required for production of the reporter protein. These labeled subunits could be inserted as pre-made products, along with the reporter gene with altered promoter (to bind to the target transcription factor), into cells with production of YOR205C and knockout YOR205C -/- cells. According to the hypothesis, YOR205Cp is involved in ribosome biogenesis. This gene could interact with ribosomal subunits to cause them to come together correctly or cause one of them to become active, enabling it to join to the rest of the ribosome. Presumably, cells with YOR205C would produce the reporter, barring any steric effects from the extra components attached to the ribosome, while cells lacking the gene would not produce the reporter protein, if YOR205C aids in the formation of and interacts with the mitochondrial ribosome.
This experiment, however, comes with a few problems. First, YOR205C may not be involved in the structural formation of the mitochondrial ribosome at all, in which case the experiment would be useless. Second, because the ribosome has a very specific shape, the transcription factor fragments added to each subunit could disrupt this shape, preventing ribosomal formation due to experimental design. Third, YOR205C may be involved in the steps of ribosome biogenesis before the pieces begin to come together; for example, it could involve post-translational modification of one of the subunits directly after translation. In this case, the absence of YOR205C would not affect how these proteins come together and would involve protein processing, causing no production of reporter gene in either cell group. To combat this problem, one could try inserting the coding sequence for a TF fragment into each DNA segment coding for the ribosomal subunits, but this method presents additional problems with control of insertion and with modification of necessary segments of each ribosomal subunit. Because of these problems, the above method might not be able to definitively illuminate the function of YOR205C in ribosome biogenesis but instead could help narrow its function down to a smaller range of possibilities.
The final hypothesis, whether or not the YOR205 protein binds to RNA and acts as an RNA helicase, could be tested using another modified version of Y2H. If transcription factor fragments can be attached to proteins and other substances (such as methyl groups) can be attached to RNA, it could be possible to use an RNA molecule with a specific sequence as a "bait" molecule in a Y2H experiment. One could then use YOR205Cp as the prey protein; addition to both into yeast cells along with binding between the two would cause transcription factor completion and production of a promoter gene. Unfortunately, if YOR205Cp is an RNA helicase, then it could bind to a specific sequence of RNA. Not only is it unknown what this sequence is, but it is unknown how long the sequence would be! Luckily, sequence analysis and comparison could be helpful in this case, since YOR205 was homologous to other RNA helicases in yeast as well as other organisms (see previous page). If the RNA binding sites of these orthologs and paralogs could be determined, they could provide a starting point for determining the correct RNA binding sequence used by YOR205Cp, as long as the RNA-binding segment of both the non-annotated gene and its homolog were conserved. If this sequence can be hypothesized, this method could not only determine whether YOR205Cp binds to RNA, but also the length and sequence information for the RNA segment to which the enzyme binds.
By performing these three experiments, one could test all three facets of the working hypothesis concerning YOR205C.
References
Campbell, A. Malcolm, and Laurie J. Heyer. Discovering Genomics, Proteomics, and Bioinformatics, 2nd ed. San Francisco and others: CSHL press, 2006.
ExPASy. Saccharomyces cerevisiae SWISS-2DPAGE Viewer. Last Updated 26 October 2006. <http://ca.expasy.org/swiss-2dpage/viewer>. Accessed 16 November 2006.
Kumar, A., Cheung, K.-H., Ross-Macdonald, P., Coelho, P.S.R., Miller, P., and Snyder, M. (2000). TRIPLES: a Database of Gene Function in S. cerevisiae. Nucleic Acids Res. 28, 81-84.
NCBI Protein Database. "NP_014840." <http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&val=99030932>. Accessed 14 November 2006.
SGD (Saccharomyces Genome Database) MCA1 gene. Updated 2006. <http://db.yeastgenome.org/cgi-bin/locus.pl?locus=MCA1>. Accessed 4 October 2006.
SGD (Saccharomyces Genome Database) YOR205C gene. Updated 2006. <http://db.yeastgenome.org/cgi-bin/locus.pl?locus=YOR205C>. Accessed 17 November 2006.
SGD glossary. 2006. <http://www.yeastgenome.org/help/glossary.html>. Accessed 16 November 2006.
Database of Interacting Proteins. UCLA. Last Updated 7 February 2004. <http://dip.doe-mbi.ucla.edu/dip/Main.cgi>. Accessed 16 November 2006.
Questions? Comments? Email brhenschen "at" davidson.edu
© 2006 Department of Biology, Davidson College, Davidson, NC 28035