|
CD4 T Cell Roles and Responses to HIV Infection |
|
This web page was produced as an assignment for an undergraduate course at Davidson College |
|
CD4 T cells activate B cells, natural killer cells, and phagocytes upon recognition of foreign antigen on Major Histology Complex class II (MHC class II), and for this reason play a pivotal role in the immune response of healthy individuals (Finberg et al., 1990). The CD4 T cell activates these effector cells by secreting cytokines; primarily interleukin-2 (IL-2) and interferon-gamma (IFN-gamma) (Stine, 1999). Activated B cells produce antigen specific neutralizing antibodies that bind to the foreign virus and prevent fusion with the cell membrane (Janeway et al., 1999 and Parren et al., 1997). |
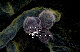
| Figure 10- This is a scanning electron microscope image (magnified 500,000x) of a HIV virion infecting a CD4 T cell. The is image is used with permission from MediaMD.com <www.cmsp.com/datav1/cg010001.htm>. |
| Natural Killer cells are antigen nonspecific lymphocytes that are activated to destroy viruses (Stine, 1999). Specifically the FcgammaRIII receptor on natural killer cells binds to IgG1 or IgG3 subclasses which are attached to the virus (Janeway et al., 1999). Finally, the phagocytes engulf foreign particles and display antigen in the lymph nodes. These, phagocytes, known as antigen presenting cells (APC) activate B and T cells for specific antigen (Stine, 1999). |
 |
|
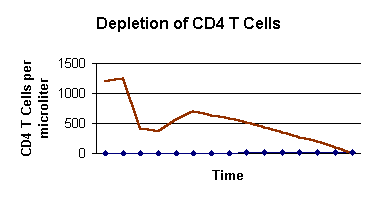
first weeks ~ mean of 10 years |
| Figure 11- The first several weeks of HIV infection are characterized by seroconversion and a drop in CD4 T cell level followed by an adaptive immune response which restores the amount of CD4 T cells to approximately 700 per microliter. The virus continues to replicated for a mean of 10 years until the patient has a CD4 T cell count below 200 per microliter. At this stage the individual has AIDS and is susceptible to many opportunistic infcetions. Adapted from Janeway et al. (1999). |
| Theories of CD4 T Cell Destruction |
| Direct Cell Killing- Infection of HIV into the CD4 T cell and subsequent budding off disrupt the cell membrane causing cell death. HIV may also disrupt internal host cell machinery resulting in cell death (Schwartz and Nair, 1999). Furthermore, bind of HIV to the CD4 T cell triggers cytotoxic CD8 T cells to attack and kill the infected cell (Janeway et al., 1999) |
| Filling of CD4 Receptor Sites- HIV may bind to the CD4 receptor on T cells in two ways. First, the gp120 portion of the gp160 moiety may bind to the CD4 receptor. Also, the gp120 may detach from the HIV virion and bind to a CD4 receptor (Wyatt and Sodroski, 1998). Cells that have gp120 bound to the CD4 receptor are susceptible to antibody-dependent cell cytotoxcity and nonantibody-dependent cytotoxic T cell attack. Furthermore, binding to the CD4 receptor causes the T cell to loose immune function despite HIV fussing and infecting the cell (Stine, 1999) |
| Syncytia Formation- Syncytia is defined as fusion of uninfected CD4 cells with infected CD4 cells to form a multinucleated cellular complex (Stine, 1999). Several in vitro studies have found a direct correlation between the presence of syncytia and the degree of cytopathic effect by HIV, however the mechanism by which this occurs is not well understood (Groenik et al., 1993). |
| Apoptosis- Programmed cell death has been shown both in HIV infected CD4 T cells and in uninfected CD4 cells with gp120 bound. The binding of gp120 to the CD4 receptor primes the T cell and when the T cell encounters antigen on MHC class II a similar signal to that by the Fas/Fas ligand is sent (Schwartz and Nair, 1999). This stimulates a signal transduction pathway that results in stimulation of caspase-activated DNase (CAD) in the nucleus (Barr et al., 1994). Upon activation of CAD the DNA of the host cell is cleaved into approximately 200 base pairs (Janeway et al., 1999). |
| Superantigens- The typical antigen or virus is capable of evoking an immune response from less than one in every million CD4 T cells (approximately 0.01%). Superantigens, however, interact with 5% to 30% of the CD4 T cells by binding to the MHC class II or T cell receptors (Stine, 1999). This hypothesis states that HIV retroviral-encoded superantigens bind to MHC class II on antigen presenting cells (APCs), thereby inducing CD4 T cells into anergy and eventually T cell death (Janeway et al., 1999). |
| Cofactors- The cofactor hypothesis states that other infectious organisms, stress, morphine-based drug use, or malnurishment may accelerate HIV expression after initial infection (Fitzgerald, 1988 and Stine, 1999; ). Newly discovered human herpes virus (HHV-6, -7, and -8) may act as cofactors in HIV infection . Moreover, cytomegalovirus and the Epstein-Barr virus have been correlated with increased HIV expression (Schwartz and Nair, 1999). |
| Impact of CD4 T Cell Destruction |
|
A healthy individual has approximately 800 to 1200 CD4 T cells per microliter, however that number is reduced in the latent stage of HIV infection until the number of CD4 T cells reaches 200 per microliter. At this stage an individual has AIDS and is susceptible to numerous opportunistic infections (Schwartz and Nair, 1999). The infection of CD4 T cells by HIV is central to the success of this virus due to the fact that CD4 is used to activate numerous immune system cells. CD4 T cells contribute the following (Stine, 1999): 1. Activate and induce macrophages and natural killer cells 2. Control maturation and induction of B cells 3. Secretion of growth and differentiation factors of lymphoid cells 4. Secretion of hematopoietic colony stimulating factors 5. Secretion of factors that induce non-lymphoid cell funtion.
|
| Home Page | Introduction | HIV Background and Structure |
| HIV Live Cycle | Treatment | References |
|
|
This web site was created for an Immunology class. Please direct correspondence to jodickens@davidson.edu. Last Updated April 21, 2000 |
