Structure of CD59
CD59 is a membrane-bound protein that is present in many types of cells and binds to homologous complement factors C8 and C9 of the terminal complement system. This small glycoprotein is made of 77 amino acids and has a molecular weight of 18- to 25-kD (McKusick, 2001). As well, CD59 is a protein that has numerous synonymous names including: protectin, H19, membrane inhibitor of reactive lysis (MIRL), P-18, homologous restriction factor-20 (HRF-20), 1F-5Ag, and the membrane attack complex-inhibitory factor (MACIF) (Messer et al., 2006). This protein can be found in numerous animals, but specifically is located on short arm of chromosome 11 in the human genome. The gene encoding CD59 has been found specifically assigned to chromosome 11p13 and 11p14 (McKusick, 2001).
The CD59 protein is presumably consisted of 4 exons. The first exon is untranslated, the second encodes the hydrophobic leader sequence of the protein, the third exon encodes the N-terminal portion of the mature protein, and the fourth exon encodes the remainder of the mature protein including the glycosylphosphatidylinositol (GPI) anchor attachment in the plasma membrane. The structure of the CD59 protein is very similar to the Ly6 family (McKusick, 2001). Similarities between the two proteins, CD59 and Ly6, are significant because they may both be involved in lymphocyte activation and differentiation at critical times. CD59 and Ly6 proteins both have ten cysteine residues placed in similar positions. As well, they are about the same size and are both GPI-anchored molecules. Last, both proteins are composed of 4 exons where the first exon is untranslated and each have following exon splices in comparable locations along their DNA (Petranka et. al., 1992). While these two proteins are similar, there are a few qualities to distinguish the two. Petranka et. al. shows evidence that Ly6 is not present on murine erythrocytes (red blood cells) , while CD59 is present on human erythrocytes (1992). Furthermore, the CD59 genes are not inducible by interferon-gamma and Ly6 genes are, however, inducible (Vakeva, 1995).
Below is the CD59 protein structure. This structure is characterized as a single cysteine-rich domain, having a hydrophobic core with three loops and a small fourth loop, which is helical (Yu et. al., 1997). The helical loop is made of two anti-parallel beta-sheets and the other three loops are three anti-parallel beta-sheets, which form the core. Connecting each of these loops are five disulphide-bonded cysteine pairs (Instructions to see these bonds are in the Figure 1 caption). As a three-dimensional structure, CD59 looks flat and disked-shaped, with a surface area of 32 nm2 (Farkas et. al., 2002).
Figure 1. The structure of the CD59 protein. The three helical loop structures, creating the hydrophobic core of CD59, are shown in yellow and the helical loop is in pink. To see the disulphide-bonded cysteine pairs - right click on the JMOL figure, select Render, Disulphide Bonds, and On. To make the disulphide bonds more visible - right click, Render, Disulphide Bonds, and 0.30 A. You can also choose to color the disulphide bonds another color to see them more clearly within the CD59 structure. Courtesy of Protein Data Bank.
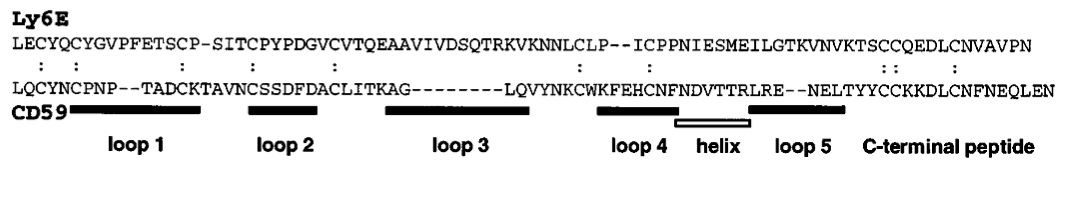
Figure 2. CD59 and Ly6 proteins have comparable sequences shown above. The conserved cysteine residues are shown by a (:). Information also includes the amino acids which form the surface loops and helix structure of CD59. Permission pending from the Journal of Experimental Medicine.
Another significant part of the CD59 molecule is its glycophosphoinosital (GPI) anchor which attaches the protein to the membrane of different cells. Without these GPI anchor proteins, such as CD59, terminal complement molecules would bind to the target cell and lyse the cell through the membrane attack complex (MAC). The presence of GPI-linked molecules also allows for lipid rafts to form, made of CD59 proteins (Cross, 2004). These anchors on CD59 proteins are simply the parts that enable these proteins to bind to cell membranes, which are essential for CD59’s function (Besa, 2005). The GPI anchor is shown in green within the membrane in Figure 5 below.
Function of CD59
The CD59 protein is a complement system regulator that can inhibit the formation of the membrane attack complex (MAC). It has also been suggested that a less common function of CD59 can influence the proliferation capacity of T cells and their ability to produce cytokines, which can influence how T cells respond to a given antigen that may enter the bloodstream. CD59 works in the innate immune system, but has also been implicated in the adaptive immune system (Longhi et. al., 2006).
There are many molecules and proteins that are used in collaboration to activate different pathways of the complement system. One of the pathways is the terminal cascade pathway. During the terminal complement cascade, a pathogen surfaces, a series of cleavage reactions occur, and C3 convertase results. C3 convertase is then cleaved into C3a, C3b, and terminal complement components (C5b, C6, C7, C8, and C9). These terminal components are used to make the membrane attack complex (MAC, C5b-9 complex). First, C5 releases a convertase molecule that releases C5b. Then, by more cleavage processes, C6 binds C5b, C7 binds C5b6, and C8 binds C5b67. Once C8 binds the other molecules the small complex C5b-8 is created. This smaller complex undergoes conformational changes to cause the hydrophobic domains to insert the membrane, attaching to the target cell. While these terminal components bind to the membrane, C9 molecules are induced and expose the hydrophobic site. At this time the MAC starts to form a non-selective ion channel pore structure (Farkas et. al., 2002 and Janeway et. al., 2005). The C5b-9 complex is said to contain up to 18 C9 molecules, which are polymerized to make this barrel-like shape (Vakeva, 1995). Once multiple C9 molecules have bound together with the other terminal components, the pore structure’s diameter of the MAC induces cell lysis of the target cell by breeching the cell’s membrane (Janeway et. al., 2005).
The protein, CD59, limits the number of C9 molecules that can interact and insert into the C5b-9 complex, inhibiting the production of the MAC. This CD59 protein inhibits the terminal pathway by blocking the formation of the membrane attack complex (MAC) by binding the complement factors, C8 and C9, within the C5b-9 complex. When CD59 binds this complex it prevents unfolding and polymerization of the final C9 pore (Farkas et. al., 2002 and Longhi et. al., 2006).
In addition, CD59 influences the outcome of a T cell response to a given antigen. Longhi et. al. has proven through research that direct interaciton between CD59 on a T cell and a specific receptor on an antigen presenting cell (APC) results in an inhibitory signal being transmitted to both the T cell or the APC. Due to this inhibitory signal, down-modulation of APC activity and consequently T cell activity results as well. Longhi et. al. also concludes that CD59 on T cells may reduce the strength of the positive signal transduction pathway delivered through the T cell receptor (TCR). The CD59 surface protein can undergo cross linking on lymphocytes to stimulate the T cell, cause organization of the cytoskeleton, and enhance proliferation. The GPI anchors on the CD59 protein are responsible for this clustering in a lipid raft type manner (Longhi et. al., 2006). A diagram for CD59 interacting with T cells and APCs is shown below in Figure 4.

Figure 3. The membrane attack complex (MAC) is made by poly C9 (in green) binding to the C5b678 complex (multicolored). CD59 is the orange protein inhibiting C9s from binding to the C5b678 complex to prevent cell lysis. Courtesy of Immunology, 6th edition, Garland Science 2005.
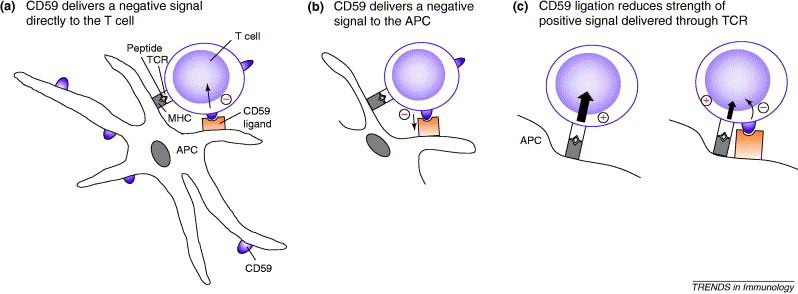
Figure 4. CD59 has an effect on T cell-APC interactions. T cells interaction with a specific receptor of an APC results in an inhibitory signal between the two cells, which reduces the strength of positive signals delivered through the TCR. Permission pending from Trends in Immunology journal.
The active site - How CD59 interacts with complement components
The complement system is designed to rid the body of dangerous pathogens. The terminal complement system kills pathogenic cells by lysing them through the MAC; however, the MAC is not always successful and can form its pore structure on a host cell and kill it rather than the pathogen. The protein, CD59, is responsible for inhibiting the C5b-9 complex formation. This way the ion channel does not form, the cell does not become lysed, and host cells will continue to live.
According to Yu et. al., it has been suggested through experimentation that CD59 specifically binds only to C9 through its active site, which later inhibits the pore formation of the MAC. The active site is suspected to be located in the vicinity of a hydrophobic groove from amino acid 16-57 (shown in the diagram below in yellow). This section has been found to be the most conserved area of the protein and a mutation of this area can disrupt its function (Yu et. al., 1997). Similarly, Kieffer et al. concludes in an experiment that the functional active site for CD59 is a hydrophobic strip composed of Tyrosine and Phenylalanine residues (1994). Yu et al. also adds that a secondary active binding site of CD59 to C8 and C9 may be on an under loop of Asn8 to Cys13. They are also conserved residues that seem to show binding, but may not be required for its inhibitory function. Finding the active site for CD59 can be beneficial to further research and design of the CD59 inhibitor to possibly block the CD59 active site so lysis may be enhanced on tumor cells, for example (Yu et. al., 1997).
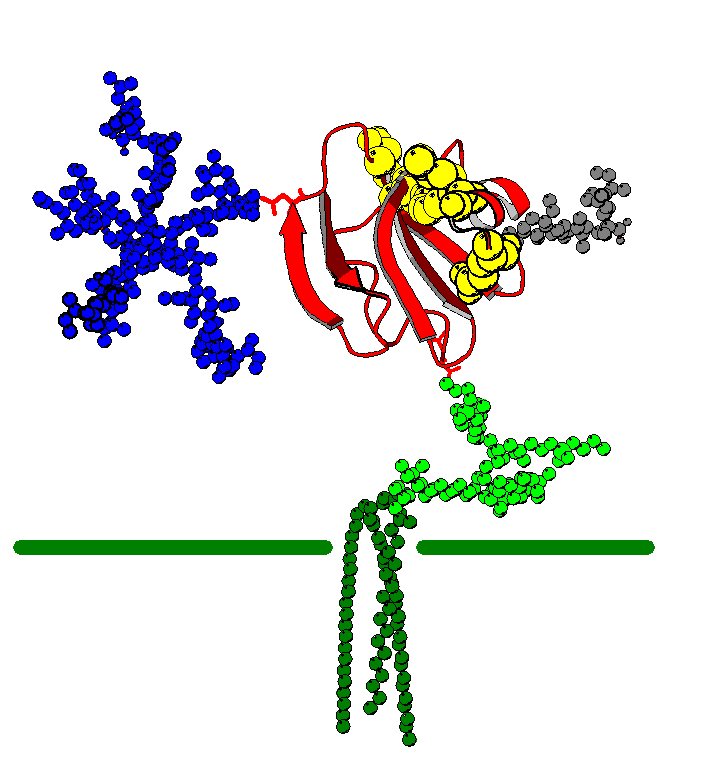
Figure 5. The active site of CD59 is show in this structure of CD59 in yellow. CD59 is attached to its GPI anchor (in green) and a N-glycosylation site (in blue). Courtesy of the Oxford Glycobiology Institute.
Where CD59 is located in the human body
CD59 can be found throughout the body as a regulator of cell lysis through the complement system on erythrocytes (red blood cells), leukocytes (white blood cells), fibroblasts, and various epithelial surfaces including the pancreas, epidermis, bronchi, kidney, and salivary glands (Vakeva, 1995). CD59 has also been found on sperm. Cross proves that CD59 is restricted to the external layer of the plasma membrane because it can be easily removed by an extracellular phophatidyl inositol-specific phospholipase C. Like GM1, CD59 was found present uniformly over the whole uncapacitated sperm surface (Cross, 2004). The presence of CD59 on sperm allows for protection from anti-sperm antibodies that may develop in the female reproductive tract during the attack of the complement system, this way sperm can react with eggs successfully (Cross, 2004). CD59 tends to be a surface protein that has increased expression when the complement system is attacking a target cell (Vakeva, 1995). While CD59 seems to be located in numerous locations in the human body, it has not been detected on B lymphocytes.
CD59 is also found in other animals. Some of the animals that have undergone CD59 analysis are mice, pigs, dogs, monkeys, horses, cows, rabbits and snakes. The strains within the different animals all have some similarities and seem to have some evolutionary conservation among their structures as well (Messer, 2006).
Deficiencies involving CD59 and its effects
CD59 is a regulatory protein for the terminal complement system to prevent cell lysis; however, the system does not always work appropriately and can therefore cause damage. A disease called Paroxysmal Nocturnal Hemoglobinuria (PNH) is a rare disease resulting from unusual susceptibility of red blood cells to the lytic action of complement. It is considered an acquired chronic hemolytic anemia, causing recurrent thrombotic events. In this disease, CD59 does not attach to the red blood cells due to somatic mutation in a hematopoietic stem cell. These mutated hematopoitic cells result in death of red blood cells by undergoing cell lysis (hemolysis) (PNH, 2006). The mutation can cause defects within the GPI anchor and create a problem keeping the CD59 protein on the cell’s surface (McKusick, 2001). PNH blood cells deficient in the GPI anchor lack membrane proteins (CD59) linked via the anchor and are degraded in the endoplasmic reticulum before being presented on the surface (PNH, 2006).
Due to this deficiency involving CD59, PNH patients suffer from various syndromes and symptoms. If the patients present the presence of anemia, then symptoms include dark colored urine, elevated reticulocyte count, and serum lactic acid dehydrogenase (LDH) levels. If anemic, patients typically lose iron in urine and can cause damage to their bone marrow as well. Another symptom involves thrombosis of the venous system, which can result in severe pain of the abdominal area, liver, skin, and brain. Last, symptoms can include spasms, aplastic episodes, and increased heart risks (Besa, 2005).
The only treatment suggestions for PNH currently are bone marrow transplants, immunosuppressive therapy, and retroviral gene therapy with CD59 (PNH, 2006 and McKusick, 2001).
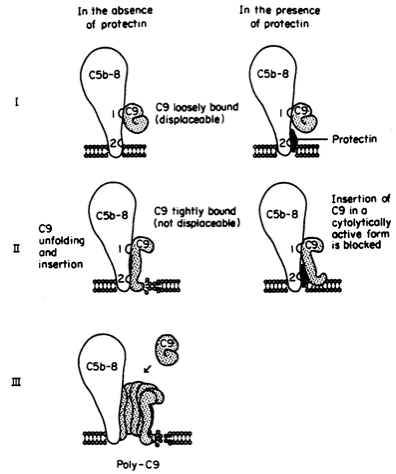
Figure 6 . Another figure demonstrating how CD59 binds to the C5b-8 complex and C9s, to prevent construction of the complete C5b-9 complex (MAC). CD59 blocks C9 from attaching to C5b-8. In PNH, CD59 is absent (left column) and the MAC forms, causing lysis of erythrocytes (RBCs). Permission pending from SUNY Stony Brook Pathology Department.
Additionally, CD59 has been found correlated with diabetes. Diabetes is a disease due to an increased level in glucose, which results in glycation and impairment of protein function. Glycation is when a sugar molecule binds a protein or a lipid molecule without the control of an enzyme. So, recently it was tested if glycation could inhibit CD59 and it was proven true. The presence of glycation in CD59 loses its MAC-inhibitory function and develops vascular proliferavative complications like diabetes (McKusick, 2001). Davies et. al. also found that loss of functional CD59 in hyperglycaemics made them more susceptible to lysis by complement. The data of experiments performed by Davies et. al. suggests that glycation-induced inactivation of CD59 is a factor contributing to anaemia in type I diabetes. It is possible that glycation near residue K41 and W40, two highly conserved amino acids, are essential to CD59 function and when they are altered, CD59 is inhibited (Davies et. al., 2004).
How CD59 can help in Xenotransplantation
In the last few years, scientists have used the human gene associated with CD59 in xenotransplants (animal organs used human transplants). One concern with transplants is that the host cells will reject the new cells, especially when a different species’ organs are transplanted. One way to help in the accepting of an animal’s organ is to use a transgenic pig or baboon with the human form of CD59, along with CD46, CD55, and DAF. By using these complement-inhibitory proteins, the human complement system will avoid damaging the pig organs and accept the new organ as its own. Transgenic pigs for the previously listed proteins are necessary for xenotransplantations or the human will undergo rejection and could suffer death (Lambigts et. al., 1998).
Resources
Besa, Emmanuel C. 4 Aug 2005. Parozysmal Nocturnal Hemoglobinuria. emedicine. Accessed 13 Mar 2006. http://www.emedicine.com/med/topic2696.htm.
Cross, Nicholas L. 10 June 2004. Reorganization of Lipid Rafts During Capacitation of Human Sperm. Biology of Reproduction. 71(4): 1367-1373.
Davies, Catherine S.; Harris, Claire L.; Morgan, B. Paul. 9 Aug 2004. Glycation of CD59 impairs complement regulation on erythrocytes from diabetic subjects. Immunology. 114: 280-286.
Farkas, Imre; Baranyi, Lajos; Ishikawa, Yasushige; Okada, Noriko; Bohata, Csaba; Budai, Denes; Fukuda, Atsuo; Imai, Masaki. 2002. CD59 bocks not only the insertion of C9 into MAC but inhibits ion channel formation by homologous C5b-8 as well as C5b-9. Journal of Physiology. 539(2): 537-545.
Kieffer, B.; Driscoll, P.C.; Campbell I.D. 1994. Three Dimensional Solution Structure of the Extracellular Region of the Complement Regulatory Protein CD59, a New Cell-surface Protein Domain Related to Snake Venom Neurotoxins. Biochemistry. 33: 4471-4482.
Lambrigts, Denis; Sachs, David H.; Cooper, David K.C. 15 Sept 1998. Discordant Organ Xenotransplantation in Primates: World Experience and Current Status. Transplantation. 66(5): 547-561.
Longhi, M. Paula; Harris, Claire L.; Morgan, B. Paul; Gallimore, Awen. Feb 2006. Holding T cells in check – a new role for complement regulators? Trends in Immunology. 27(2):102-108.
Messer, Jeff; Vargas, Guillermo; Murillo, Andres. Analysis of Protectin. Accessed 9 Mar 2006. http://www.astarte.csustan.edu.
McKusick, Victor A. 9 Oct 2001. CD59 Antigen, Protectin. Online Mendelian Inheritance in Man (OMIM). Accessed 9 Mar 2006 http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=OMIM.
Paroxysmal Nocturnal Hemoglobinaria (PNH). SUNY Stony Brook Pathology Department. New York State Univeristy. 2006. Accessed 12 Mar 2006. http://www.path.sunysb.edu.
Petranka, J.G.; Fleenor, D.E.; Sykes, K.; Kaufman, R.E.; Rosse, W.F. 1992. Structure of the CD59-encoding gene: further evidence of a relationship to murine lymphocyte antigen Ly-6 protein. Proceedings of the National Academy of Sciences. 89: 7876-7879.
Protein Data Bank (PDB). Structure keyword: 1CDR. Desposited 01 June 1994. Accessed 15 Mar 2006. http://www.rcsb.org/pdb/.
Vakeva, Antti. 1995. Complement Activation in Myocardial Infarction. Academic Dissertation. University of Helsinki, Finland. http://www.helsinki.fi/~vakeva/thesis.html.
Wormald, M. No date. Oxford Glycobiology Institute. Molecular Model Archives. Accessed 8 Mar 2006. http://www.bioch.ox.ac.uk/glycob/.
Yu, By Jinghua; Abagyan, Ruben; Dong, Shanghong; Gilbert, Alexander; Nussenzweig, Victor; Tomlinson, Stephen. 17 Feb 1997. Mapping the Active Site of CD59. Journal of Experimental Medicine. 185(4): 745-753.

Davidson College Immunology Home Page
© Copyright 2005 Department of Biology, Davidson College, Davidson, NC 28036