
CD1: Antigen Presentation and T Cell Function
Summary
This website describes the important characteristics and immunological significance of the antigen presenting molecule CD1. CD1 is structurally and functionally similar to MHC class I and II molecules; however, CD1 presents lipids rather than polypeptides on antigen presenting cells. CD1 has also evolved a unique path of intracellular trafficking, processing, and loading of lipid antigens. CD1 presentation of lipid antigens can elicit an immune response from a subclass of T cells aptly named Cd1-restricted T cells. Because much of the significance of the CD1 molecule lies in its ability to stimulate the unique effector and helper functions of the Cd1-restricted T cell, I will also describe this subpopulation of T cells. Throughout my discussion of CD1, I will be making many comparisons to MHC class I and II molecules. Below is a link that describes these molecules.
MHC Class I and II Antigen Presenting Molecules
Genes
Despite its functional similarity to MHC class I and II molecules, the CD1 genes are not located within or linked to the Major Histocompatibility Complex but are located in a 170,000bp region of chromosome 1q23.1 in humans (Brigl and Brenner, 2004). The following five CD1 genes have been identified in humans: CD1A, CD1B, CD1C, CD1D, and CD1E (Brigl and Brenner, 2004). Brigl and Brenner (2004) use lowercase letters to correspond to the proteins encoded by each of these genes, and I will adapt this nomenclature for this website (i.e. CD1A genes encode protein CD1a). While humans have five different distinct genes of CD1, the murine genome contains only a pair of CD1D orthologs (Brigl and Brenner, 2004). It is significant to note that homologs for human CD1A, CD1B, CD1C, and CD1E are absent in the murine genome because much of information about the functionality of CD1, especially as it relates to autoimmunity and protection from disease, has been gathered from murine experimental models. In contrast to the highly variant MHC class I and II genes, limited polymorphism has been observed in CD1 genes (Han et. al, 1999). Brigl and Brenner (2004) speculate that the limited polymorphism observed in CD1 genes may be due to the limited diversity of lipid tails that CD1 binds and displays.
Figure 1 Genomic map of the human and mouse CD1 loci. Five distinct CD1 isoform encoding genes exist on human chromosome 1 while the mouse only contains two orthologs of CD1D. A possible chromosomal translocation may have led to the deletion of the other isoform genes (Brigl and Brenner, 2004). Figure taken from Brigl and Brenner (2004), permission pending. |
Structure
Like the MHC class I complex, “CD1 polypeptides are expressed as heterodimers composed of the CD1 heavy chain noncovalently paired with b2-microglobulin” (Brigl and Brenner, 2004). Studies suggest that the CD1 molecule can exist in the absence of b2-microglobulin, but the significance of these rare b2-microglobulin-free forms is unknown(Brigl and Brenner, 2004). The atomic structure of CD1 resembles that of MHC class I molecules; however, the binding groove of CD1 is significantly larger than both MHC class I or II molecules (Brigl and Brenner, 2004).The antigen binding grooves also differ between CD1a,b,c, and d suggesting that the different isoforms may present structurally diverse lipid-containing antigens (Brigl and Brenner, 2004).
Figure 1. Jmol image of CD1 mouse antigen presenting molecule (CDB Protein Database 1cd1) |
Antigen Binding Capabilities
Studies to date suggest that CD1 molecules present only lipid components of antigens (Brigl and Brenner, 2004). CD1 molecules are also believed to present self antigens and, therefore, may play a role in interaction and selection of certain T cells (Brigl and Brenner, 2004). α-Galactosylceramide(aGalCer) is an example of a glycosphingolipid believed to be presented by CD1 and has been used to study activation of CD1-restricted T cells (Brigl and Brenner, 2004).
Presentation of CD1
The process of presentation of functional CD1 is not fully understood, but some components of the process have been identified. The first step in intracellular trafficking of CD1 to the surface is newly synthesized CD1 molecules are assembled with b-2 microglobulin in the ER with the help of chaperone proteins (Brigl and Brenner, 2004). The manner in which CD1 binds lipids is not fully understood, but it is believed that some type of cellular lipid initially blocks binding of inapropriate antigens, a function analogous to that of the invariant chain in MHC class II assembly (Brigl and Brenner, 2004). This CD-1/"prelipid" complex is delivered from the Golgi body to the plasma membrane via the secretory pathway. CD1 is then reinternalized where it binds a foreign lipid or another self lipid (see Figure 3) (Brigl and Brenner, 2004).


| Figure 3. Comparison of intracellular trafficking of CD1 and MHC Class I and II molecules. Figure taken from Brigl and Brenner (2004), permission pending. |
Expression of CD1 on Antigen Presenting Cells
CD1 molecules are primarily expressed on dendritic cells and other antigen presenting cells including B cells (Brigl and Brenner, 2004). CD1 is predominantly expressed by cortical thymocytes and interdigitating cells. CD1a, b, c, and d are expressed in different frequencies on different types of antigen presenting cells. Langerhans' cells are CD1a positive while mantle cells and some of the peripheral blood and spleen B cells express CD1c (Brigl and Brenner, 2004). Dermal dendritic cells and interlocked dendritic cells in lymph nodes tend to be predominantly CD1b positive (Brigl and Brenner, 2004). Follicular denditic cells, monocytes, macrophages and all postthymic neoplasms are CD1 negative (Naeim). Low levels of CD1d can be detected on most monocytes, macrophages, dendritic cells, B cells, and some nonlyphoid cells (Brigl and Brenner, 2004). Interestingly, CD1d can be detected in high concentration on B cells in the mantle zone, but only low levels of CD1d can be found in germinal centers (Exley et. al, 2000). While CD1a, b, and c tend to be expressed in higher concentrations than CD1d on antigen presenting cells, CD1d is found on a much broader range of cells (Brigl and Brenner, 2004).
Unique Maturation of CD1+ Antigen Presenting Cells
Unlike MHC class II molecules which increase their concentration sharply during maturation, levels of CD1 molecules seem to stay constant or elevate only slightly during dendritic cell maturation (Brigl and Brenner, 2004). This suggests that the process of CD1 trafficking is induced by dendritic cell maturation as is the case in MHC class II molecules (Brigl and Brenner, 2004). Figure 4 shows the expression of surface CD1 and other important cell surface molecules during myleoid dendritic cell maturation (Brigl and Brenner, 2004).

Figure 4. Cell surface receptor molecule expression in maturation and differentiation of CD1-restricted T cells. Figure taken from Brigl and Brenner (2004), permission pending. |
T Cell Recognition of CD1
The T cell receptor(TCR) of CD1-restricted T cells interacts with foreign, self, and synthetic lipids presented by CD1 with fine specificity (Brigl and Brenner, 2004). Differences of only a few carbon molecules in a lipid tail presented by CD1 has been shown to greatly effect bioactivity of CD1-restricted T cells (Brigl and Brenner, 2004). CD1 is able to bind to a wide variety of lipids, examples of which are shown in Figure 3. However, recent murine experiments suggest that CD1-d restricted T cells may have the capacity to recognize foreign microbial antigens in the absense of TCR-mediated recognition, suggesting their possible significance in early microbial infection (Brigl and Brenner, 2004).
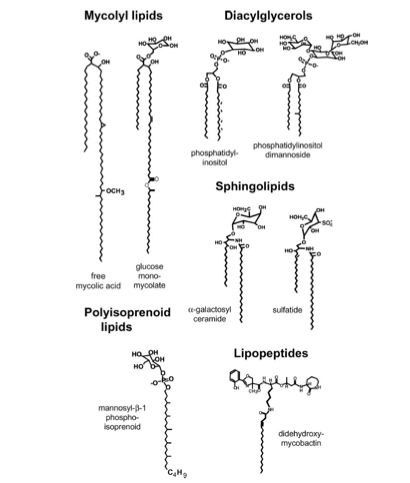
Figure 5. Examples of lipids presented by CD1. Figure taken from Brigl and Brenner (2004), permission pending. |
CD1-restricted T cells
The lipid antigen presenting molecule CD1 stimulates a repertoire of unique CD1-restricted T cells. These CD1-restricted T cells appear to go through processes of negative and positive selection in the thymus similar to MHC restricted T cells (Brigl and Brenner, 2004). Some CD1-reactive T cells have been found to possess the coreceptors CD4 or CD8 while other CD1-restricted T cells have been foudn to be double negative for the CD4 and CD8 coreceptors (Brigl and Brenner, 2004). CD1-restricted T cells also seem to be isoform sensitive in a fashion analogous to MHC restriction (i.e. CD1a binds only to a specific CD1a sensitive TCR) (Brigl and Brenner, 2004). Relationships between isoform specific TCRs and costimulatory molecules are unclear. Experiments with murine models which posses only CD1d suggest that CD1-restricted T cells recirculate through peripheral tissues and enter lymph nodes through afferent lymphatics in contrast to their MHC restricted counterparts which use primarily high endothelial venules (Brigl and Brenner, 2004). Effector functions of CD1-restricted T cells seem to be diverse. CD1 has been found to stimulate both Th1 and Th2 effector functions (Brigl and Brenner, 2004). CD1-restricted T cells have been found to promptly release cytokines upon activation by CD1 and costimulatory molecules (Brigl and Brenner, 2004). Many of these cytokines play an important role in recruitment and activation of other immune cells. CD1-restricted T cells have also been shown to perform cytolytic functions upon activation (Brigl and Brenner, 2004). Activated CD1-restricted T cells then will either undergo apoptosis or undergo extensive proliferation (Brigl and Brenner, 2004). It is not fully understood what causes some CD1-restricted T cells to persist and multiply and others to undergo apoptosis.
Antimicrobial and antitumor activities of CD-1
One must recognize that the antimicrobial activity of CD1 is indirect because effector CD1-restricted T cells must be present for an appropriate immune response. CD1 seems to play a significant role in immune responses to infectious diseases and some types of uncontrolled cell growth (i.e. cancer) (Brigl and Brenner, 2004). In humans, CD1 a, b, and c have been associated with effective immune responses to leprosy and mycobacterial infections (Brigl and Brenner, 2004). CD1-restricted T cells constituted the primary CD8+ T cell response to M. Bovis BCG immunization (Brigl and Brenner). Studies have also shown CD1 presentation to be downregulated following exposure to live M. tuberculosis. Interestingly, dead M. tuberculosis did not elicit the same downregulattion of CD1 (Brigl and Brenner, 2004). Brigl and Brenner(2004) believe that these studies may point to CD1 downregulation as a possible immune system evasion strategy of pathogens (Brigl and Brenner, 2004). Studies also point to possible roles of CD1-restricted self reactive T cells in providing “dendritic cell instruction”(possibly directing migration to lymph nodes) which may help bridge the gap between the innate and adaptive immune response (Brigl and Brenner, 2004). Low levels of CD1d are believed to be present at functional levels on tissue macrophages and resting monocytes at all times which suggests that CD1d and CD1d-restricted T cells may play a role in early response to pathogens. Studies suggest that CD1 antigen presentation occuring during microbial infection may stimulate CD1-restricted T cells which contribute to Th1 biased antimicrobial immune responses (Brigl and Brenner, 2004). Studies of human tumors have shown that tumor infiltrating dendritic cells often present CD1, suggesting that CD1 may present tumor glycolipid antigens (Brigl and Brenner, 2004). These findings suggest that CD1 expression could become of prognostic value for certain cancers (Brigl and Brenner, 2004). CD1a has been shown to be expressed by the main cells in several cancer-like conditions including Histiocytosis X, a malignancy of Langerhans cells (Brigl and Brenner, 2004). Together, these studies suggest that CD1 participates in "an extensive array of host defense and immunolopathologic processes" (Brigl and Brenner, 2004). Murine experimental models have expanded our knowledge of possible roles of CD1 in desease and cancer immune responses.
CD1d and the murine immune system
Manipulation of murine models has shown CD1-d contributes to host immune responses in bacterial, parasitic, viral, and fungal infections (Brigl and Brenner, 2004). CD1d deficient mice have shown decreased resistance to Pseudomonas aeruginosa and Borrelia bugdorferi (Brigl and Brenner, 2004). Activation of macrophages and recruitment of neutrophiles seemed to be impaired in CD1d deficient mice infected with Pseudomonas aeruginosa while CD1d deficient mice infected with Borrelia bugdorferi showed impaired antibody production (Brigl and Brenner). CD1d and CD1d-restricted T cells appear to participate in immune defense against M. tuberculosis and M. bovis BCG, but their specific role is not clear (Brigl and Brenner, 2004). While most murine experiments suggest CD1d and CD1d-restricted T cells are beneficial to bacterial immune responses, there appear to be exceptions. Listeria monocytogenes seems to benefit from CD1d expression as host survival increased with anti-CD1d mAB blocking (Brigl and Brenner, 2004). CD1d-restricted T cell activation may also increase tisssue damage (Brigl and Brenner, 2004). A variety of experiments testing parasitic immune responses suggest CD1-restricted T cells play an active role in antiparasitic responses through increasing protective Th2 immunity (Brigl and Brenner, 2004). A subset of CD1d-restricted T cells which use TCRα-invariant as well as diverse γδTCRs seem to participate in immune responses to viruses (Brigl and Brenner, 2004). CD1d-restricted T cells have also been found to contribute to tumor rejection in murine models; however, the method of CD1 directed tumor suppression is not yet known (Brigl and Brenner, 2004).
αGalCer and immunosurveillance by CD1d-restricted T cells
αGalCer, a glycosphingolipid found in marine sponges, has been shown to stimulate CD1-restricted T cells in humans and mice. αGalCer and similiar compounds "may have therapeutic potential for the treatment of infectious diseases and the improvement of vaccine efficacy" (Brigl and Brenner, 2004). Studies suggest that αGalCer stimulates antitumor responses against a number of different tumors and different levels of metastasis in mice (Brigl and Brenner, 2004). The anti-tumor activity of αGalCer has been implicated in the following tumors: melanomas, colon, lung, prostate, breast and renal cell carcinomas, and lymphomas (Brigl and Brenner, 2004). In these scenarios, CD1d-restricted T cells seem to stimulate, probably through cytokine secretion, NK cells and CD8+ cytotoxic T cells which then perform the direct antitumor effector functions (Brigl and Brenner, 2004). Recognition of antigens presented by CD1 also seems to be critical in the mediation of CD1-restricted tumor immunosurveillance (See Figure 6) (Brigl and Brenner, 2004). CD1-restricted T cells have also been shown to suppress antitumor immunity through inhibiting the development of protective anti-tumor responses (Brigl and Brenner, 2004). The seemingly parodoxical roles of CD1d/CD1d-restricted T cells illustrates how little we really know about the role of CD1 in the immune system.

Figure 6. Proposed model of CD1/CD1d-restricted T cell function in natural tumor immunosurveilance. The top process shows CD1d-restricted T cell mediated tumor rejection while the bottom shows CD1d T cell suppression of antitumor immunity. Note that NKT represents CD1-restricted T cells. Figure taken from Brigl and Brenner (2004), permission pending. |
CD1d-restricted T Cells and autoimmunity
Nonobese diabetic(NOD) mice, a mouse strain which develops a form of autoimmune diabetes similar to type 1 diabetes in humans, has been shown to have a deficiency of CD1d-restricted T cells (Brigl and Brenner, 2004). Increasing levels of CD1d-restricted T cells in these NOD mice was shown to alleviate the condition leading researchers to believe CD1-restricted T cells play an important role in protecting against this autoimmune condition (Brigl and Brenner, 2004). Type 1 diabetes may be prevented by CD1d-restricted T cells which stop the differentiation of autoreactive T cells into effector cells (see Figure 7) (Brigl and Brenner, 2004). CD1d-restricted T cells also have been studied in relation to Experimental Autoimmune Encephalomyelitis (EAE), a murine model of Multiple Sclerosis (Brigl and Brenner, 2004). "Manipulations that enhance the Th2 bias of CD1d-restricted T cells strikingly reduce the development of EAE" (Brigl and brenner, 2004).
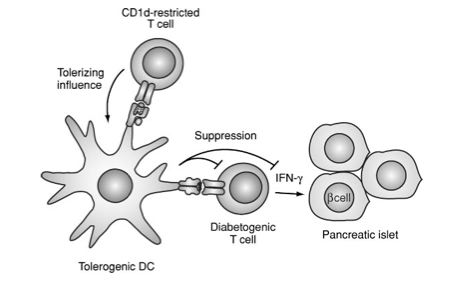
Figure 7. Proposed method of protection conferred by CD1-d-restricted T cells. Figure taken from Brigl and Brenner (2004), permission pending. |
Brigl, Manfred and Michael Brenner. 2004. CD1: Antigen presentation and T cell function. Annual Review of Immunology 22, 817-890.
Exley M., Garcia J., Wilson, S.B., Spada, F. Gerdes, D., et. al. 2000. CD1d structure and regulation on human thymocytes, peripheral blood T cells, B cells, and monocytes. Immunology 100, 37-47.
Han, M., Hannick, L., DiBrino, M. and M.A. Robinson. 1999. Polymorphism of human CD1 genes. Tissue Antigens 54, 122-127.
Naeim, Faramarz. Clusters of Differentiation Antigens. Department of Pathology. UCLA School of Medicine. 1996. Accessed 12 Mar 2006. http://www.pathology.ucla.edu/educ/lecture/pathrev/antigen/cdfront.htm
Protein Data Bank (PDB). Structure keyword: 1CD1. Desposited 02 April 1997. Accessed 14 Mar 2006. http://www.rcsb.org/pdb/
Acknoledgements:
Special thanks to Dr. Manfred Brigl and Dr. Michael Brenner of Brigham and Women's Hospital and Hardvard Medical School whose review article CD1:Antien Presentation and T cell Function formed the basis of my website.
Return to Immunology Main Page
E-mail questions & comments to: jufried@davidson.edu