*This web page was produced as an assignment for an undergraduate course at Davidson College *
Introduction
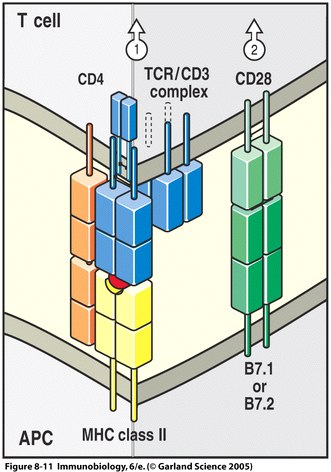
Our immune system is a complex and fascinating system that relies on numerous protein interactions and signaling capabilities in order to protect our bodies from foreign invaders. Many of these pathways require more than one signal before the cascade can begin to ensure that the appropriate response to a given circumstance can be provided. One of the best characterized co-stimulatory pathways involves the B7 molecules during the activation of T cells by antigen-presenting cells (APCs). The first signal for T cell activation is provided by the binding of a naïve T cell through its receptor to its specific peptide:MHC complex on an APC. Before T cell activation can be complete, however, a second, co-stimulatory signal must be provided. This is the role of the B7 molecules, found on cells that activate T cells. When a T cell binds to the peptide:MHC complex, the B7 molecules bind to their ligand, CD28, which is found on the surface of resting and activated T cells. This second signal is what allows the T cell to be fully activated, allowing the immune response to continue on its way to ridding the body of foreign invaders (Epstein et al., 1996).
Figure 1. Two signals are necessary for complete T cell activation. The first is provided by the binding of the T cell receptor to its specific peptide:MHC molecule. The co-stimulatory signal is provided by the binding of a B7 molecule on the antigen presenting cell (APC) to its ligand, CD28, on the T cell (Janeway et al., 2005).
Structure
There are two B7 molecules, B7.1 (also called CD80) and B7.2 (CD86), both of which are members of the immunoglobulin superfamily, a collection of proteins that all have one or more immunoglobulin domains or immunoglobulin-like domains. Both B7.1 and B7.2 are homodimers, and each have an extracellular V-like domain and C-like domain. They each also have a transmembrane anchor and a short cytoplasmic tail. The two molecules have very similar structures, although B7.2 is slightly larger with 304 amino acids rather than the 254 amino acids of B7.1. The regions of the B7 molecules used for binding to their ligands also differs; their extracellular domains have only 27% amino acids in common (Freeman et al., 1998).
Expression
The B7 molecules are primarily found on cells that activate T cells, namely activated dendritic cells, activated B cells, and activated macrophages. The B7 molecules are also found constitutively on memory B cells. In addition to the T cell activating cells, B7.2 is also expressed at low levels on unstimulated dendritic cells and granulocytes, while B.1 is expressed on fibroblasts. The reasons for B7 expression on cells such as granulocytes and fibroblasts is unknown, but may indicate the possibility of other roles of the B7 molecules other than T cell co-stimulation (Epstein et al., 1996).
Dendritic cells are the most effective stimulators of T cell activation. Although it has been shown that low levels of B7.2 molecules are present on immature dendritic cells, these cells are not yet ready for T cell activation. Typically once an immature dendritic cell (such as a Langerhans’ cell) engulfs foreign material, it travels to the lymph nodes where it begins expression of the B7 molecules (both B7.1 and B7.2). This expression is necessary for the complete activation of T cells that will eventually be activated by the dendritic cell (Janeway et al., 2005).
Macrophages are another type antigen presenting cell with the capacity for activating naive T cells. Macrophages do not begin B7 expression, however, until they have engulfed material that is identified as foreign. More specifically, it is the interferon-γ involved in macrophage activation that induces the expression of B7 molecules on the macrophage surface (Janeway et al., 2005).
B cells are similar to dendritic cells and macrophages with respect to B7 molecules in that they do not express B7.1 or B7.2 until foreign matter is recognized by its receptors. The memory B cells with continued lifespan after an infection is cleared up express the B7 molecules constitutively on their surfaces to allow for quick activation of T cells in the event of another attack by the same foreign antigen (Freeman et al., 1998).
There are several warning signs that these APCs are specifically reacting to. These include bacterial lipopolysaccharides, bacterial DNA, and Toll. These are typically what induce the initial production and expression of the B7 molecules on T cell-stimulating cells. Once the B7 molecules have begun expression on the cell surfaces, several cytokines contribute to their continued expression. These include BM-CSF in dendritic cells, IFN- γ in macrophages, and IL-7 in B cells (Freeman et al., 1998).
Function
As it has been previously stated, the B7 molecules (when bound to their CD28 ligand of the T cell) provide the co-stimulatory signal necessary for T cell activation. When a naive T cell binds to its specific peptide:MHC complex, the CD28 on the T cell's surface binds to the B7 molecules on the antigen presenting cell. These two signals together lead to the production of several cytokines. The importance of B7 co-stimulation is evident in incomplete T cell stimulation when anti-B7 molecules are present (Epstein et al., 1996).
An extremely important effect of B7:CD28 signaling is the synthesis of interleukin-2 (IL-2). IL-2 is the cytokine primarily responsible for the proliferation and differentiation of activated T cells. When a T cell comes into contact with an APC presenting its specific antigen:MHC complex without co-stimulation by the B7:CD28 complex, the cell does not produce sufficient IL-2 to allow for T cell proliferation and differentiation, therefore preventing an appropriate immune response . When co-stimulation by the B7 molecules is present with the antigen:MHC complex on the APC, however, the co-stimulation pathway leads to stabilized IL-2 mRNA and more IL-2 production and secretion. Cytokine expression is carefully controlled, and one mechanism for this is the unstable nature of cytokine mRNA. The B7:CD28 signaling allows for more stable IL-2 mRNA. In addition to this, the co-stimulatory pathway also activates the transcription factors responsible for the transcription of IL-2 mRNA (Janeway et al., 2005). B7 transfectants have also been shown to co-stimulate production of the IL-2 receptors on the activated T cell's surface. To further promote T cell proliferation, the B7:CD28 signaling pathway also induces production of anti-apoptotic proteins (Freeman et al., 1998).
Once a T cell is activated, the B7:CD28 pathway can work with other signaling pathways through a dialogue such that each pathway aids in the continuation of the other. One example of this is the CD40:CD40L (ligand) pathway. CD40 is a protein found on APCs, and its ligand is found on activated T cells. While the CD40:CD40L pathway stimulates the expression of more B7 molecules on the APC and more CD28 on the T cell, the B7:CD28 pathway activates the T cell to express more CD40L and the APC to express more CD40 (Janeway et al., 2005). This is important because the CD40:CD40L pathway plays an important role in T cell effector function once T cells are completely differentiated (Freeman et al., 1998). This relationship is supported by a study that showed when CD28 signaling is blocked, the CD40 pathway is not enough to allow priming by the APC (Prilliman et al., 2002). This circle of activation allows both pathways to continue signaling to the T cell.
Regulation
The drastic changes that come with T cell activation require that the process be tightly regulated. Signaling through the B7 molecules, then, must be efficiently controlled. This regulation is carried out by a second ligand of the B7 molecules, CTLA-4 (CD152). The expression of CTLA-4 on T cell surfaces only begins after the T cell's activation. CTLA-4 molecules bind roughly 20 times more tightly than CD28 to the B7 molecules. This means that as T cell activation continues, more CTLA-4 is being expressed on the T cell surface (Janeway et al., 2005). Although CTLA-4 expression never gets to be as dense on the T cell surface as does CD28 (only about 2-5% of CD28 expression level), the much higher avidity for the B7 molecules effectively decreases the strength of the B7:CD28 signaling pathway. One model of this decrease suggests that when high amounts of B7 molecules are being expressed following activation of the T cell, both the CD28 and CTLA-4 ligands are binding the B7 molecules. This model says that once B7 expression starts to decrease, the vast majority of the remaining surface B7 molecules are bound by CTLA-4 because of its higher avidity for B7 (Freeman et al., 1998). In this way, the B7:CD28 signals are controlled and production of IL-2 within the activated T cell is maintained at an appropriate level.
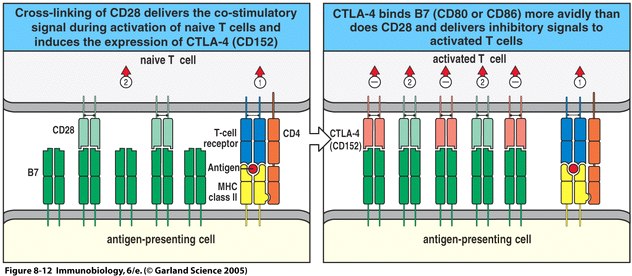
Figure 2. CTLA-4 regulates B7:CD28 signaling. The co-stimulatory signal allows complete activation of the T cell, as seen on the left. Once activation begins, CTLA-4 is expressed on the activated T cell and begins to bind available B7 molecules with more avidity than the CD28 molecules on the T cell surface. The B7:CD28 signaling pathway therefore decreases due to the limited available B7 molecules available for CD28 binding (Janeway et al., 2005).
B7 molecules and Autoimmunity
It is important to note what happens to T cells that do not receive the necessary co-stimulation through the B7:CD28 pathway. Such cells enter a state of angery, meaning the T cell becomes eternally unresponsive to further stimulation, even if co-stimulation is present in a second encounter with its specific antigen. In other words, these T cells will never be able to become activated. This effect combined with the necessity of both T cell activating signals coming from the same cell has to do with avoiding autoimmunity. If a T cell recognized self tissues as its specific antigen and then received a co-stimulatory signal from a neighboring cell, the T cell would become activated and autoimmunity would occur (Janeway et al., 2005).
The requirement of co-stimulatory signals greatly reduces the chance that T cells will become activated and attack self antigens. Even some nonbacterial antigens do not provide the co-stimulatory signal required for complete T cell activation and adjuvants must be used to aid the immune attack. This use of adjuvants illustrates the important requirements and therefore crucial role of B7 co-stimulatory molecules in immune responses involving T cells (Janeway et al., 2005).
Promising studies
Much research is currently being conducted in the hopes that new information about the co-stimulatory function of the B7 molecules may provide insight into possible cures for certain diseases. Some of this research deals with the possibility of using B7-transfected cells as a way of controlling tumor growth. One study looked at the effects of providing B7 co-stimulation to mice with mammary tumors expressing human epithelian mucin (MUC1). The researchers wanted to determine the reason for weak immogenicity against the tumor cells. The study showed that when B7.1 or B7.2 was expressed with the MUC1, the mice not only survived for longer but also went for longer periods of time before any tumors were detected (Vasilevko et al., 2003).
Another study looked at the effects of CD4 positive and CD25 positive T cells and their ability to suppress autoreactive T cells when B7 molecules are also expressed on the target T cells. Mice that did not express the B7 molecules were unable to suppress the autoreactive T cells and suffered disease. When the B7 molecules were restored on the T cells, the mice were once again able to control and suppress the T cells reacting to self antigens. This suggests possible uses for B7 molecules in dealing with autoimmune disorders (Paust, et al., 2004).
Another study looked at the effects of blocking both the B7:CD28 and CD40:CD40L pathways in an attempt to avoid immunogenicity against gene therapy adenovirus vectors. This promising work showed that blocking both B7.1 and B7.2 as well as CD40L completely suppressed humoral and cellular attacks against the vectors (Ziller et al., 2002). This shows a promising future for the role of the B7 molecules in dealing with the negative effects associated with gene therapy and the body's rejection of foreign materials.
A similar study looked at the effects of small molecules specifically chosen because of their ability to inhibit the binding of B7.1 to CD28 and CTLA-4. The study was able to identify a specific binding site on B7.1 that is not present on B7.2 or on mouse B7.1. The hope of the researchers is that this site could provide a "hot spot" for inhibiting any role B7.1 molecules have in immunogenicity against transplants (Erbe et al., 2002).
References
Epstein FH, Reiser H, Stadecker MJ, 1996. Costimulatory B7 molecules in the pathogenesis of infectious and autoimmune diseases. The New England Journal of Medicine 335:1369-1376.
Erbe DV, Wang S, Xing Y, Tobin JF, 2002. Small molecule ligands define a binding site on the immune regulatory protein B7.1. Journal of Biological Chemistry 277:7363-7368.
Freeman GJ, Boussiotis VA, Gribben JG, Sharpe AH. “B7 (CD80 and CD86).” Encyclopedia of Immunology. 2nd ed. 1998.
Janeway, C.A., Travers, P., Walport, M., Schlomchik, M. Immunobiology 6th Ed: The Immune System in Health and Disease. New York: Garland Publishing, 2005.
Paust S, Lu L, McCarty N, Cantor H, 2004. Engagement of B7 on effector T cells by regulatory T cells prevents autoimmune disease. Proceedings of the National Academy of Sciences of the United States of America 101:10398-10403.
Prilliman KR, Lemmens EE, Palioungas G, Wolfe TG, Allison JP, Sharpe AH, Schoenberger SP, 2002. A crucial role for B7-CD28 in transmitting T help from APC to CTL. Journal of Immunology 169:4094-4097.
Vasilevko V, Ghochikyan A, Sadzikava N, petrushina I, Tran M, Cohen EP, Kesslak PJ, Cribbs DH, Nicolson GL, Agadjanyan MG, 2003. Immunization with a vaccine that combines the expression of MUC1 and B7 co-stimulatory molecules prolongs the survival of mice and delays the appearance of mouse mammary tumors. Clinical & Experimental Metastasis 20:489-498.
Ziller C, Stoeckel F, Boon L, Haegel-Kronberger H, 2002. Transient blocking of both B7.1 (CD80) and B7.2 (CD86) in addition to CD40-CD40L interaction fully abrogates the immune response following systemic injection of adenovirus vector. Gene Therapy 9:537-546.
Questions or comments? jeking@davidson.edu
Return to immunology home page