*This page was created as an assignment for an undergraduate class at Davidson College
The role of E and P-selectin in Leukocyte
Recruitment to Sites of Infection
Brief Overview
Selectins are a family of cell surface adhesion molecules that are produced as a part of the inflammatory response of innate immunity. This website will focus primarily on the functions of two members of the-selectin family, E and P-selectin. E and P-selectin are produced by the endothelial cells that line the vessel walls of the circulatory system. They function to recruit lymphocytes to sites of inflammation, which are indicative of the presence of pathogen. E and P-selectin are very important to our body’s immune system because they aid in acute immune responses by directing lymphocytes to specific areas of infection and are utilized by both the innate and adaptive immune systems.
How it works
The role of E and P-selectin begins with the inflammatory immune response. Macrophages are activated when they encounter certain toxins that causes them to release cytokines (Janeway et. al. 2005). These cytokines cause inflammation to occur in the surrounding tissues which takes the form of pain due to lymphocytes leaking into tissues and the chemicals they release, redness and heat due to increased blood flow to the area, and swelling due to the increased permeability of blood capillaries.
One of the cytokines released by macrophages is Tumor Necrosis Factor α (TNFα). TNFα stimulates the endothelial cells lining blood vessels to express the surface adhesion molecule P-selectin (Janeway et. al. 2005). Within a couple hours a second surface adhesion molecule, E-selectin, is produced. Together E and P-selectin slow the motion of leukocytes through the bloodstream by causing them to roll along the endothelial surface, allowing other molecules to interact with the slowed leukocytes to stop them and promote their movement into the tissues. E and P-selectin expression can also be stimulated by interleukin-1 (IL-1) and lipopolysaccharide (LPS) (Leeuwenberg et. al. 1992). LPS is a component of the membranes of many gram negative bacteria. LPS binding proteins on macrophages can stimulate them to produce inflammatory cytokines when they come in contact with their ligand (Janeway et. al. 2005).
Leukocyte Rolling and Extravasation
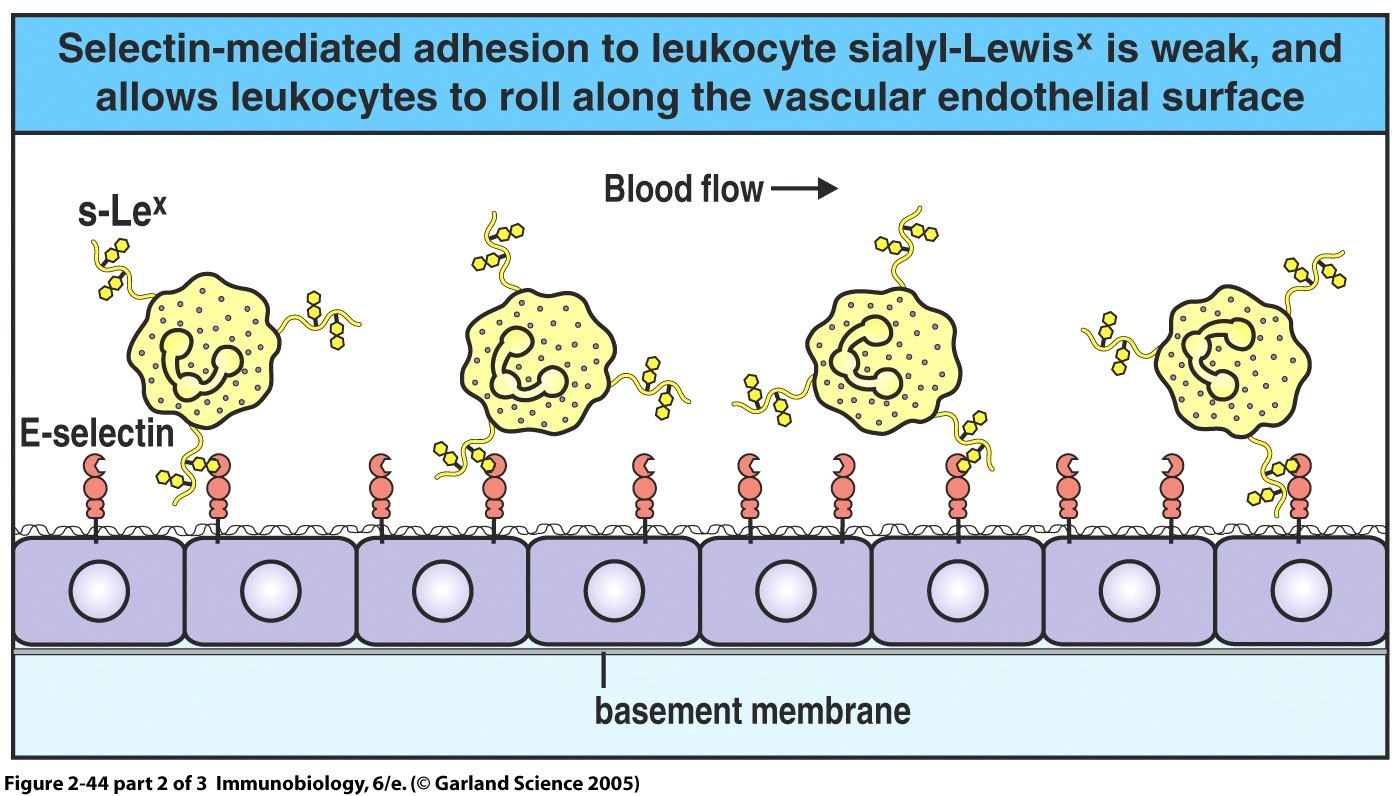
Figure 1 shows leukocyte rolling along the surface of endothelial cells. E-selectin is binding reversably with carbohydrate on the leukocyte cellular membrane. P-selectin is not shown. (From Janeway et. al.)
Both E and P-selectin are known to bind with Sialyl Lewis x (sLex)-like glycans which are expressed in relatively high numbers by circulating leukocytes (Zou et. al. 2005). Furthermore, P-selectin also binds to P-selectin glycoprotein ligand-1 (PSGL-1) which is modestly expressed on human leukocytes (Borges et. al. 1997).
A 1995 study by Steegmaler et. al. has also revealed an E-selectin specific ligand, which they labeled ESL-1. ESL-1 is structurally similar to a recently discovered chicken cysteine-rich fibroblast growth-factor receptor except for a 70 amino acid long chain at the amino-terminal domain of the ESL-1 ligand. A separate study, published in 2005 by Zou et al., suggests that E-selectin can also bind to the P-selectin ligand, PSGL-1. In this study, microspheres were either conjugated with sLex or PSGL-1, and were tested for their adherence to endothelial cells expressing E-selectin. They found that the initial rate of adhesion to the endothelial cells was greater in the microspheres that expressed PSGL-1 even though the microspheres that expressed sLex had greater ligand densities on their cell surfaces.
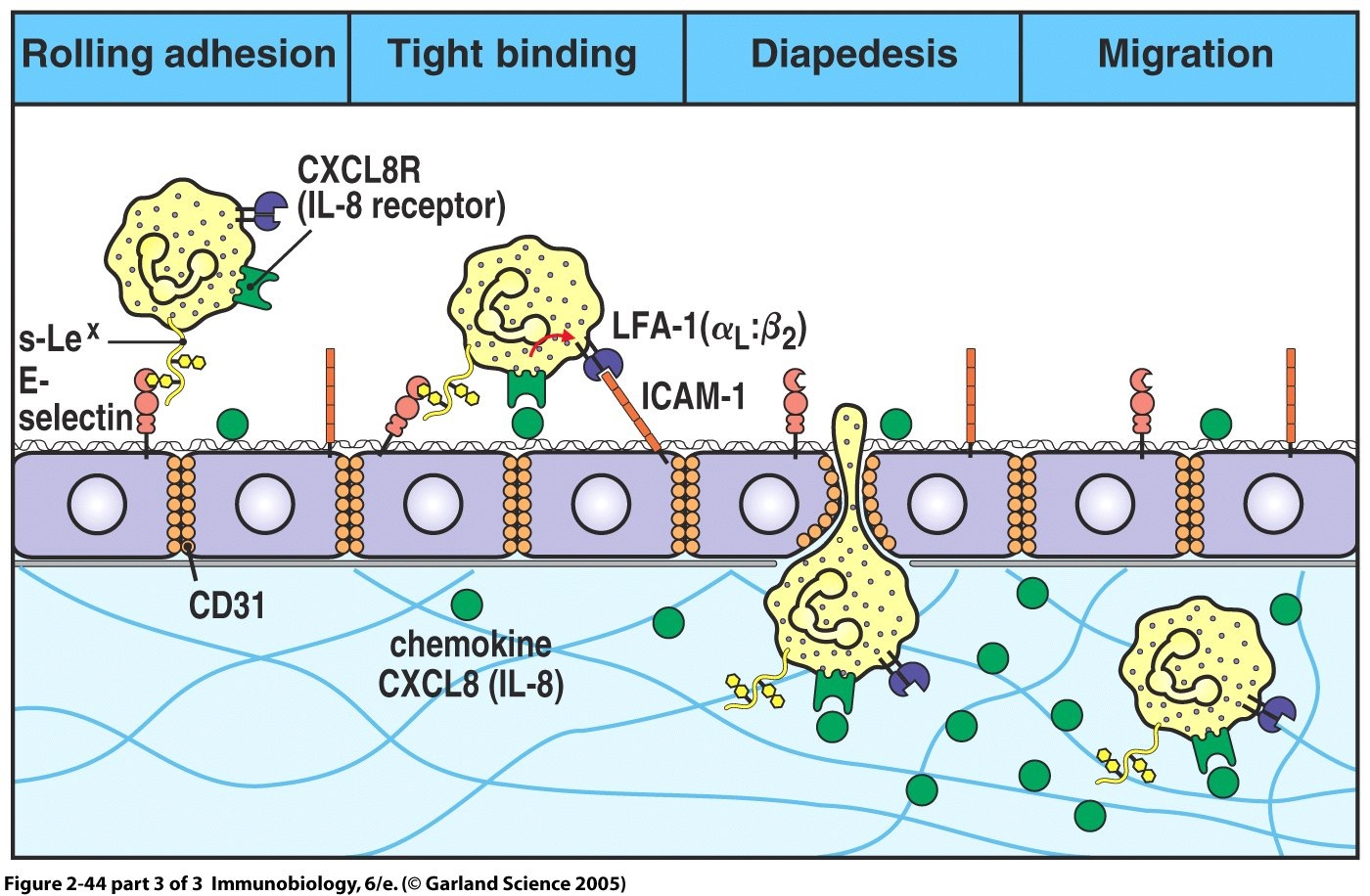
Leukocyte rolling occurs due to the brief, reversible binding of E and P-selectin with their complementary molecules, which are expressed on the surface of passing leukocytes. Leukocyte recruitment by E and P-selectin is analogous to throwing a tennis ball at a Velcro surface. An individual Velcro hook will have little effect on the motion of the ball. It will clasp onto the surface and quickly be released. However, the combined adhesive effect of many Velcro hooks will cause the ball to roll and eventually slow it down. Tight adhesion to the rolling leukocyte is performed by another molecule, intercellular adhesion molecule 1 (ICAM- 1) which binds to the integrins LFA-1 and CR3 on the leukocyte surface and arrests the motion of the rolling leukocyte. Stopping the leukocyte allows it to enter the tissues by secreting proteases to breach the endothelial basement membrane, a process known as diapedesis.
Innate and Adaptive Immune Responses
The selectin family of cell surface adhesion molecules is critical to cell recruitment in the innate and adaptive immune responses. The recruitment of neutrophils to sites of inflammation is a part of the innate immune response because it is a non-specific recruitment that occurs for every inflammatory response. Work published in 1992 by Von Andrian et. al. suggests that a third selectin molecule, L-selectin, is responsible for the recruitment of neutrophils. They found that neutrophil rolling was strongly reduced by antibody inhibition or enzymatic removal of L-selectin. These experiments involved the obstruction of L-selectin binding sites by the inclusion of receptor-specific antibodies and/or the enzymatic cleaving of L-selectin to obstruct its function. Neutrophils were unable to recognize or adhere to endothelial cells in the absence of L-selectin even though these cells expressed the surface integrins LFA-1 and CR3. A separate experiment by Mattila et. al. (2005) found that activation through a G protein associated stimulus or ligation of L-selectin causes L-selectin to cluster, causing an increase in its adhesiveness.
Selectins are also involved in the adaptive immune response with the recruitment of activated lymphocytes to sites of inflammation. One of their remarkable features is the ability to recruit T helper 1 (Th1) cells and exclude T helper 2 (Th2) cells. Austrup et. al. (1997) demonstrated that Th1 cells could be blocked from migrating into inflamed skin by P and E-selectin-specific antibodies. They concluded that this was evidence that there must be some sort of receptor-associated mechanism involving E and P-selectin that allows endothelial cells to discriminate between Th1 and Th2 cells. One month later a separate report published by Borges et. al. (1997) determined that Th1 cells bind to P-selectin by means of PSGL-1, and P-selectin binding was completely inhibited by PSGL-1 specific antibodies. However, both Th1 and Th2 cells express PSGL-1. Borges et. al. addressed this issue by suggesting that Th1 and Th2 cells possess different forms of PSGL-1, and they designated Th1 cells as having the “binding form of PSGL-1.”
E-selectin versus P-selectin
P-selectin is kept near the cell surface of endothelial cells, on the side that is exposed to the blood vessel lumen, in vesicles called Weibel-Palade bodies (Janeway et. al. 2005). The triggering of TNFα cell surface receptors on endothelial cells causes these vesicles to merge with the cell membrane, resulting in the expression of P selection on the surface of endothelial cells within minutes of cytokine recognition. E-selectin, on the other hand, is not stored in the cell and has to be transcribed, translated, and transported to the cell surface. The production of E-selectin is stimulated by the expression of P-selectin, and it takes about two hours, after cytokine recognition, for E-selectin to be expressed on the endothelial cell’s surface, with maximal expression of E-selectin occurring around 6-12 hours after cytokine recognition by the cell (Leeuwenberg et. al. 1992). E-selectin production levels-out 24 hours after the cytokine signal, and continues to be expressed for three days.
Dermal and Pulmonary Inflammatory Disease in Double and Triple-Selectin-Null Mice
An experiment by Collins et. al. (2001) sought to evaluate the roles of E, P, and L-selectin in leukocyte rolling. They did this by introducing null mutations into stem cells to produce mice that lacked E and P-selectin. The same process was also used to produce mice that lacked E, P, and L-selectin. Surprisingly, E/P double mutant mice developed skin and pulmonary disease, but E/L/P triple-mutant mice did not. Collins et. al. concluded that the absence of L-selectin in E/P mutants alters their inflammatory response. However, little is known about the specific role of L-selectin in leukocyte recruitment (Mattila et. al. 2005).
Wrap-UP
Leukocyte adhesive molecules are 15 times more adhesive to inflamed endothelium than non-inflamed endothelium, 6 times more adhesive to cytokine inflamed endothelium than non-cytokine-treated endothelium, and 10 times more adhesive to trauma-induced inflamed endothelium than non-trauma-induced endothelium (Sakhalkar et. al. 2003). We know that cytokines cause the expression of E and P-selectin by inflamed endothelium which leads to the recruitment of leukocytes to those tissues, but the exact details of their functions have yet to be discovered and continue to be tested through experimental science.
References
Austrup F, Vestweber D, Borges E, Lohning M, Brauer R, Herz U, Renz H, Hallmann R,Scheffold A, Radbruch A, Hamann A. 1997. P- and E-selectin mediate recruitment of T-helper-1 but not T-helper-2 cells into inflamed tissues. Nature 385:81-83 <http://www.nature.com/cgi-taf/DynaPage.taf?file=/nature/journal/v385/n6611/abs/385081a0.html> Accessed 2006 Mar 16.
Borges E, Tietz W, Steegmaier M, Moll T, Hallmann R, Hamann A, Vestweber D. 1997. P-selectin glycoprotein ligand-1 (PSGL-1) on T Helper 1 but not on T Helper 2 cells binds to P-selectin and supports migration into inflamed skin. J. Exp. Med. 185(3): 573-578. <http://www.jem.org/cgi/content/full/185/3/573> Accessed 2006 Mar 16.
Collins RG, Jung U, Ramirez M, Bullard DC, Hicks MJ, Smith CW, Ley K, Beaudet AL. 2001. Dermal and pulmonary inflammatory disease in E-selectin and P-selectin double-null mice is reduced in triple-selectin-null mice. Blood 98(3):727-735. Pub Med <http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=11468173&query_hl=5&itool=pubmed_docsum> Accessed 2006 Mar 16.
Janeway C, Travers P, Walport M, Shlomchik M. 2005. Immunobiology: The Immune System in Health and Disease. New York, New York: Garland Science Publishing. p. 23, 80-84.
Leeuwenberg JF, Smeets EF, Neefjes JJ, Shaffer MA, Cinek T, Jeunhomme TM, Ahern TJ, Buurman WA. 1992. E-selectin and intercellular adhesion molecule-1 are released by activated human endothelial cells in vitro. Immunology 77(4):543-9. Pub Med <http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=1283598&query_hl=2&itool=pubmed_DocSum> Accessed 2006 Mar 16.
Mattila PE, Green CE, Schaff U. 2005. Cytoskeletal interactions regulate inducible L-selectin clustering. American Journal of Physiology 289(2):323-332.
Steegmaler M, Levinovitz A, Isenmann S, Borges E, Lenter M, Kocher HP, Kleuser B, Vestweber D. 1995. The E-selectin- ligand ESL-1 is a variant of a receptor for fibroblast growth factor. Nature 373:615-620. <http://www.nature.com/nature/journal/v373/n6515/abs/373615a0.html> Accessed 2006 Mar 16.
Von Adrian UH, Hansell P, Chambers JD, Berger EM, Torres Filho I, Butcher EC, Arfors KE. 1992. L-selectin function is required for beta2-integrin-mediated neutrophil adhesion at physiological shear rates in vivo. AJP-Heart and Circulatory Physiology 263(4): 1034-H1044. <http://ajpheart.physiology.org/cgi/content/abstract/263/4/H1034> Accessed 2006 Mar 16.
Zou X, Patil VRS, Dagia NM. 2005. PSGL-1 derived from human neutrophils is a high-efficiency ligand for endothelium-expressed E-selectin under flow. American Journal of Physiology 289(2):C415-C424.
If you have any questions about this website you can contact me at jolatting@davidson.edu
Return to My Home Page
Return to Davidson College Home Page
Return to the Davidson College Immunology Home Page