NFκB is a transcription factor, and a member of the Rel family of proteins. In humans the genes encoding for NFκB are found at locations 4q.23-34 and 11q.12-13, for p50 and 65 subunits respectively (OMIM). The protein is normally found in the cytosol, bound non-covalently to an inhibitor protein, IκB (Janeway et al. 2005). Originally discovered in B cells as the transcription factor for the κ chain in immunoglobulins, NFκB is known today to be involved in a wide variety of cell pathways including, but not exclusively, stress induced, immune, and inflamatory responses (NFkb component). All in all over 100 genes' transcriptions are promoted by NFκB (Hiscott et al. 2001). The protein is also known to be involved in the development of several cell types as well as having an active role in several diseases.
NFκB is commonly activated through a Toll pathway via TLR-4; however several variations and non-classical pathways do exist that activate NFκB. The cascade starts with a microbial substance (for this cascade assume a lipopolysaccharide from Gram-negative bacteria) binding to CD14 which then associates with TLR-4 on the surface of the cell. This association triggers the cytoplasmic domain of TLR-4 (TIR) to bind and activate adaptor protein (MyD88), which interacts with the death domain of serine/theronine innate immunity kinase (SIIK), activating SIIK. SIIK then activates TRAF6 which in turn activates IκBα and IκBβ to form a dimmer (Iκk) that phosphorylates IκB. When phosphorylated IκB then dissociates from NFκB, allowing NFκB to enter the nucleus and promote translation of genes (Janeway et al. 2005). These genes include (but are not exclusive to) IL-2, IL-2 receptor (IL-2R), IL-8, interferons, Cyclin-D1 (a protein involved in the regulation og the G1 phase of the cell cycle), and many others (Imaginex).
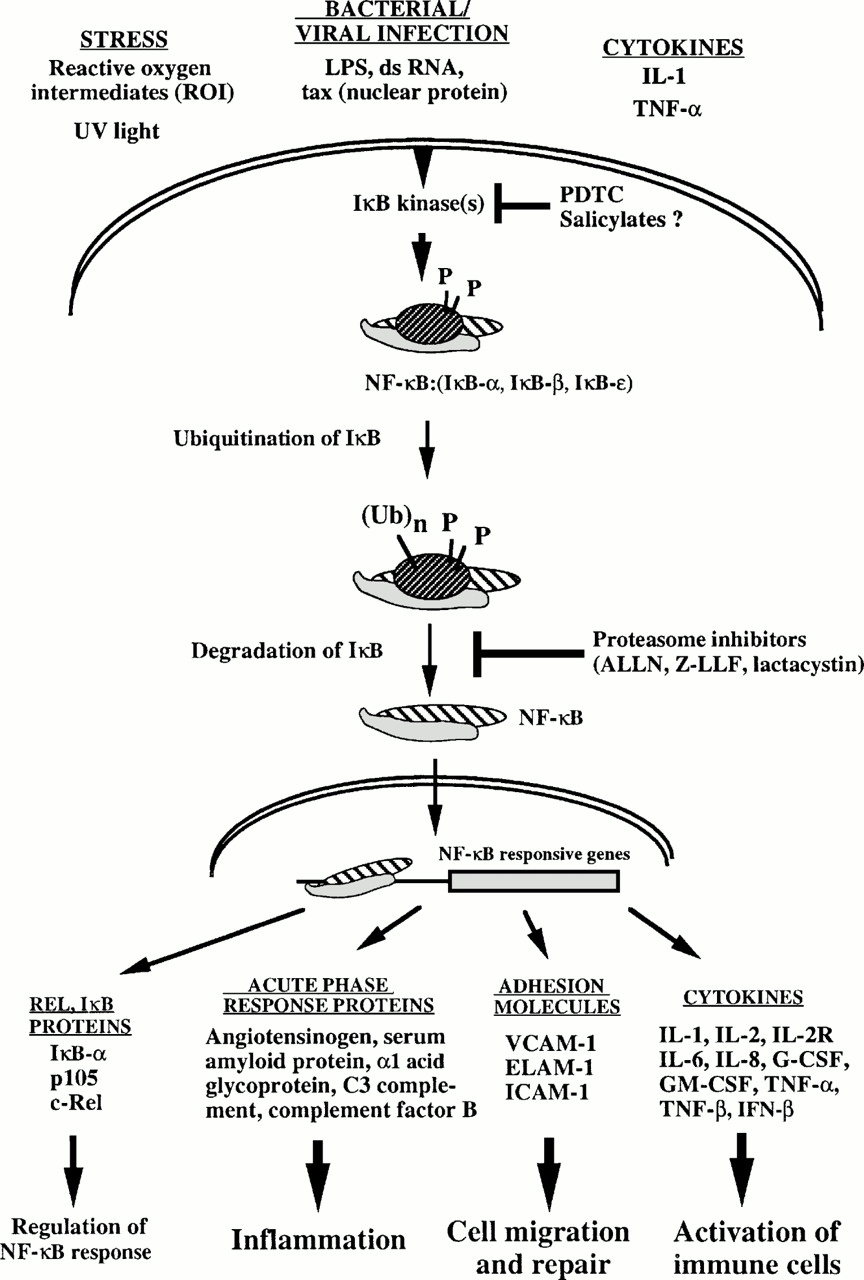
Figure.1 This image provides the general signal activation pathway for NFκB as described in the paragraph above. (McFadden 2006)
NFκB has five subunits[p50 (TNFRSF5)/p105 (NFκB1), p52/p100 (NFκB2), REL, p65/RELA and RELB]; however the p50/p65 heterodimer is the most important for most responses (Pajonk et al. 2001). p50 and p65 dimerize around a 10 base pair region called the κB site. The binding sequence of the κB site is 5'-GGGRNYYYCC-3' (R=purine, Y=pyrimidine, and N=and base) (NFkb component).
Figure 2. This is a JMOL image of NFκB bound to a κB site on a DNA molecule. Taken from the RSC PDB protein databank. (RSC PDB 1VKX)
NFκB: Clinical Studies and Information
Many studies surrounding NFκB are centered on cancer. Studies suggest that NFκB is a key component in providing the survival signal in several types of cancer cells (Pajonk et al. 2001) . Activation of NFκB promotes the production of several proteins involved in the survival signal, countering the effects of TNF (Tumor Necrosis Factor) and other death promoting proteins. NFκB has also been linked with potentially lethal damage repair (PLD repair) in cells recieving radiation therapy, allowing the cells to resist radiation-induced cell death. Inhibition of NFκB is a potentially useful tool for increasing radiosensitivity of cancerous cells (Ding et al. 2003). Normally high doses of radiation can activate NFκB, using a non-classical pathway, providing cancer cells with a stronger survival signal and more resistance to apoptosis. Inhibition of NFκB would make cells more suceptable to signals transduced by TNF, Bax, Bad and other death promoting proteins, because the survival signal that counters these signals would be weakened. Also, lower rates of PLD repair are observed in cells where NFκB is inhibited. Thus inhibition of NFκB would allow the DNA damage done by radiation to stimulate apoptisis at a greater rate (Pajonk et al. 2001).
NFκB has implications in the activation of iNOS genes. iNOS produces nitric oxide (NO), a powerful reducing agent. Reactive free radicals like NO can cause serious damage to DNA by oxidizing the DNA molecule. This type of DNA damage has been shown to lead to Alzheimer’s disease. Extensive genetic data segust that β-amyloid peptide (Aβ) plays a role in the neurodegenerative cascade of Alzheimer's disease. Aβ has also been noted to activate NFκB. In Akama's study he claims that NO production occurs through a NFκB dependent mechanism. While no mechanism for this cascade is known, Akama supports his claim by showing that iNOS promoter activity is severly depressed in cells with inactive NFκB (Akama et al. 1998).
NFκB has also been shown to play an essential role in HIV. The efficient gene replication of HIV-1 is partially attributable to the ability of the virus to sucessfully activate and bind NFκB to its gene sequence (Hiscott et al. 2001). There are a few hypothesis to how HIV activates NFκB. One study seguests that cleavage of Bcl-2 by the viral proteases leads to activation of NFκB due to the inability of cleaved Bcl-2 to maintian its normal level of supression of oxidative agents, and the oxidative sensitivity of NFκB (Strack et al. 1996). It has also been proposed that a cascade triggered by viral and cell membrane fusion involving CD4 activates NFκB. Once active the NFκB complex binds to HIV LTRs (long terminal repeats) at promoter regions, activating viral protease transcription (Hiscott et al. 2001).
There are several drugs that can affect the activity of NFκB through interactions with proteins that are involved in the cascades that regulate NFκB; however those that interact with NFκB directly do so in two common ways: directly blocking the activation of the NFκB complex, or directly blocking the duffusion of the complex. Two such drugs are 6-Amino-4-(4-phenoxyphenylethylamino)quinazoline (an inhibitor of the activation of NFκB), and NFκB SN50 (an inhibitor of the diffusion of the NFκB complex into the nucleus). 6-Amino-4-(4-phenoxyphenylethylamino)quinazoline was discovered by Tobe et. al. in 2003. Since then no literature has been published reguarding the mecanism that is used by the drug to inhibit NFκB activation (Liu 2006). NFκB SN50 inhibits diffusion of NFκB through the neuclear membrane by binding to the complex and physically inhibiting its diffusion (Puolaki et al. 2002).
Akama KT, Albanese C, Pestell RG, Van Eldik LJ. Amyloid ß-peptide Stimulates Nitric Oxide Production in Astrocytes through an NFkB-dependent Mechanism. Proceedings of the National Academy of Sciences of the United States of America May 12, 1998 Vol. 95, No. 10, 5795-5800.
Ding G., Honda N., Nakahara T., Tian F., Yoshida M., Hirose H., Miyakoshia J.. Radiosensitization by Inhibition of I?B-a Phosphorylation in Human Glioma Cells. Radiation Research 2003 Vol. 160, No. 2, 232–237.
Hiscott J., Kwon H., and Génin P.. Hostile takeovers: viral appropriation of the NF-kB pathway. Journal of Clinical Investigation January 2001, Volume 107, Number 2, 143-151
Imaginex, Innovations in Functional Genomics. NFkB Pathway. <http://www.imgenex.com/NF-kBPathway.php> Accessed 2006 Mar 15.
Janeway, A.C, P. Travers, M. Walport., J.M. Shlomchik.2005. Immunobiology: The Immune System in Health and Disease. 6th ed. New York, NY: Garland Publishing. pp 228-229.
Liu G. 2006 Mar 16. technical [Personal email]. Accessed Mar 16 2006.
McFadden J. NFkB. <http://www.surrey.ac.uk/SBMS/ACADEMICS_homepage/mcfadden_johnjoe/img/NF-kB.jpg> Accessed 2006 Mar 16
NFkb component. Enhanceosome. <http://www.biochem.umd.edu/biochem/kahn/molmachines/enhancesomes/NFkB.html> Accessed 2006 Mar 15.
[OMIM] Online Mendelian Inheritance in Man. 2006 Feb 17. NUCLEAR FACTOR KAPPA-B, SUBUNIT 1; NFKB1. <http://www.ncbi.nlm.nih.gov/entrez/dispomim.cgi?id=164011>. Accessed 2006 Mar 16.
[OMIM] Online Mendelian Inheritance in Man. 2005 Sept 19. V-REL AVIAN RETICULOENDOTHELIOSIS VIRAL ONCOGENE HOMOLOG A; RELA. <http://www.ncbi.nlm.nih.gov/entrez/dispomim.cgi?id=164011>. Accessed 2006 Mar 16.
Pajonk F., McBrideb W.. The Proteasome in Cancer Biology and Treatment. Radiation Research 2001 Vol. 156, No. 5, 447–459.
Strack PR, Frey MW, Rizzo CJ, Cordova B, George HJ, Meade R, Ho SP, Corman J, Tritch R, Korant BD. Apoptosis Mediated by HIV Protease is Preceded by Cleavage of Bcl-2. Proceedings of the National Academy of Sciences of the United States of America Sep. 3, 1996 Vol. 93, No. 18., 9571-9576.
Poulaki V, Mitsiades CS, Joussen AM, Lappas A, Kirchhof B, Mitsiades N. Constitutive Nuclear Factor-B Activity Is Crucial for Human Retinoblastoma Cell Viability. American Journal of Pathology Ded 2002 Vol. 161, No. 6, 2229-2240.