This web page was produced as an assignment for an undergraduate course at Davidson College
Lyme Disease
Introduction
Lyme disease is the most common tick-borne disorder in the northern hemisphere
characterized by inflammation of the skin, joints, nervous system and heart.
It was first identified in 1975 in the town of Old Lyme, CT after an outbreak
of juvenile rheumatoid arthritis. Lyme disease in the United States is caused
by the spirochete Borrelia burgdorferi first identified in 1982 by
Dr. Willy Burgdorfer and would have been the most serious new disease had the
AIDS virus not been identified at this time; it is also the second to AIDS as
the fastest growing disease in the world. In Europe and Asia this disease is
caused by related species, B. afzelii and B. garinii (Guerau-de-Arellano
et al. 2005).
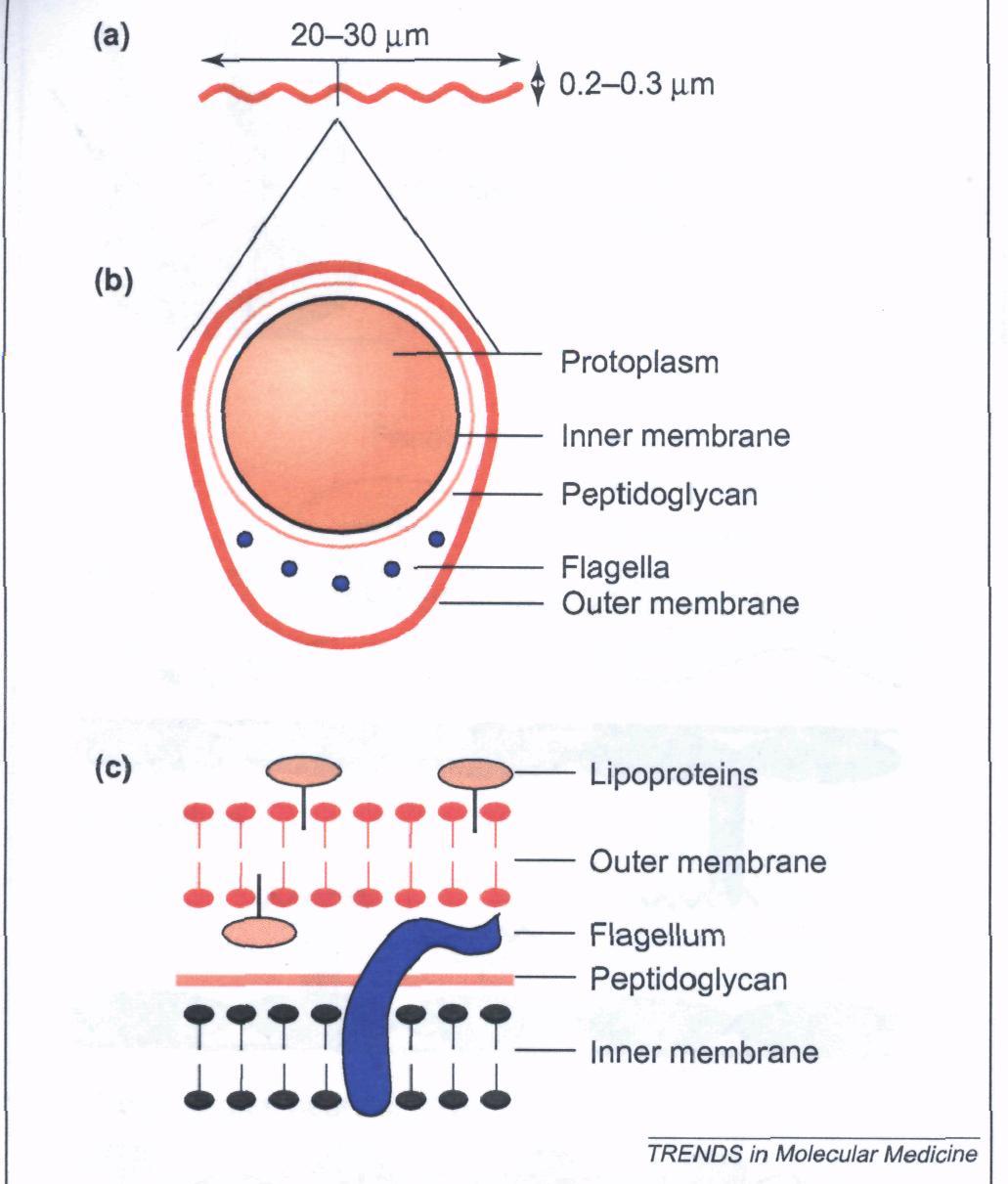
B. burgdorferi is a gram-negative bacterium with a peptidoglycan layer encasing a protoplasmic cylinder surrounded by an outer membrane and resembles a wave measuring 20-30µm in length and 0.2-0.3µm in width (Guerau-de-Arellano et al. 2005). The bacterium contains a myriad of flagella and is rich in lipoproteins, most importantly the outer-surface proteins (Osps) A-F (Schell et al. 1998; Figure 1). These Osps are encoded from the one linear chromosome the spirochete contains, consisting of nine circular and twelve linear plasmids (Guerau-de-Arellano et al. 2005). This agent is transmitted via ticks of the genus Ixodes, mostly Ixodes scapularis in North America and I. ricinus and I. persulcatus Europe and Asia; ticks must feed for more than 48 hours to transmit the Borrelia bacterium (Guerau-de-Arellano et al. 2005).
Figure 1. The Structure of B. burgdorferi. Panel A represents its general shape. Panel B represents a cross-sectional view. Panel C represetns the inner and outer membranes separated by a petpidoglycan (Permission Pending; Guerau-de-Arellano et al. 2005).
Clinical Manifestations
When a ticks feed on humans, in injects the spirochete B. burgdorferi into the skin and Lyme disease begins to develop according to three distinct stages (Figure 2). Lasting days to four weeks is the earliest stage, where the spirochete has not metastasized and the local residency causes erythema migrans, an inflammatory rash, as well as other general symptoms (Figure 3; Guerau-de-Arellano et al. 2005). This rash is generally expressed as a round circular patch resonating outward; this rash occurs in 75% of patients infected with Lyme disease and tends to resolve from the inside-out, leaving a bull’s eye mark on the skin (Frey 2002). As B. burgdorferi progresses in the body, it begins to spread through the blood and lymph and begins to invade the heart, joints and nervous system in the disseminated second stage that can lasts weeks to months. This stage is characterized by carditis, arthritis and neuropathies, with the most prevalent being arthritis. However, a correlation has been found which presents data suggesting that B. burgdorferi found in the United States is the most arthritogenic, whereas the B. afzelii and B. garnii found in Europe and Asia tend to give signs more consistent with dermatological and neuroborreliosis symptoms. The chronic late stage can last months to years if untreated and can lead to chronic forms of arthritis and neural problems (Guerau-de-Arellano et al. 2005).

Figure 2. The sages of Lyme disease and the symptoms associated with each stage (Yanagihara et al. 1997).
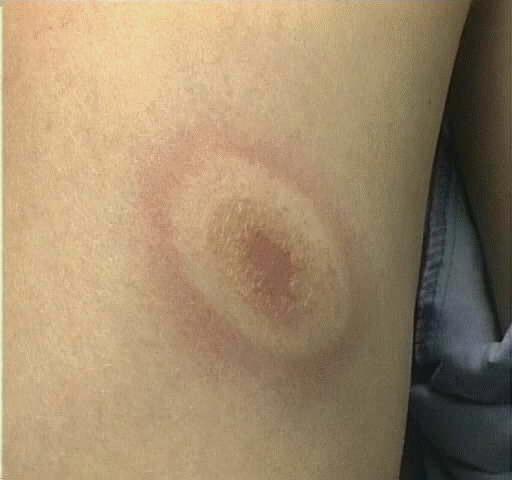
Figure 3. The erythema migrans rash associated with the early stages of Lyme disease Source:http://www.pathguy.com/lectures/infect.htm
Transmission
The spirochetes involved are well suited to survive in either the tick or a
mammalian host and expresses different proteins depending on which host it invades.
In an unfed tick, OspA is expressed at high levels, however, upon tick feeding,
OspA is down regulated and OspC is upregulated. This is because OspA allows
the bacterium to adhere to the gut of the tick by binding to the tick receptor
for OspA (TROSPA) to colonize. When a tick attaches to a mammalian host, the
changes in temperature, pH and nutrient intake cause a change in the expression
of these outer-surface proteins (Guerau-de-Arellano et al. 2005).
Most specifically, OspC is not upregulated until temperatures of approximately
32-37°C, whereas OspA is mostly found in temperatures around 24°C (Yamagihara
et al. 1997). OspC is necessary for B. burgdorferi to colonize
the human host. The exact function of OspC has not yet been characterized (Guerau-de-Arellano
et al. 2005).
Another protein essential for transmission is OspE. OspE binds to OspE-related
proteins (Erps) and complement-regulator-acquiring surface proteins (CRASPs)
to recruit compliment inhibitory proteins factor H and factor-H-like-1 to inhibit
the lysis of the antigen via the bacterial invasion; this inhibition ensures
the initial survival of the spirochete, as compliment does not become activated
(Guerau-de-Arellano et al. 2005). These CRASPs can also serve to bind
and degrade C3 convertase, an essential component of the compliment cascade,
making B. burgdorferi a compliment resistant bacterium (Bubeck-Martinez
2005).
Also important for transmission and immune evasion is the protein named VlsE.
The locus for this protein contains as many as 15 silent cassettes which allow
for changes in the conformation of the expressed protein. Antibodies produced
can not be efficiently made, as the structure of the protein continues to change.
This allows the spirochete to continue its survival as it evades the host’s
B cell immune responses (Bubeck-Martinez 2005).
B. burgdorferi can than begin to spread to the rest of the body via other expressed proteins. The OspC expressing spirochetes can bind integrins such as αIIbß3 and αVß3 expressed on platelets and endothelium, respectively. A heparin sulfate also facilitates the endothelial binding via Bgp, Borrelia glycosaminoglycan, on the bacterium. Once in the tissues, the spirochete can bind collagen with or without the proteoglycan decorin, as well as fibronectin (Guerau-de-Arellano et al. 2005; Figure 4).
![]()
Figure 4. Transmission and metastisis of B. burgdorferi and molecules involved (Permission pending; Guerau-de-Arellano et al. 2005)
Pathogenesis
Lipopolysaccharids (LPS), lipoproteins, and flagella are similar in many pathogens.
The interactions of these molecules cause inflammatory signals to be produced
via Toll-like receptors (TLRs). Innate immune cells, such as the macrophage,
bind lipoproteins of B. burgdorferi via the TLR2 receptor aided by
an LPS-binding protein (LBP) expressed during bacterial infections. Once LBP
binds, it can transfer the complex to CD14 to enable TLR2 recognition. This
binding results in the activation of NFκB, a transcription factor to upregulate
cytokines and adhesion molecules (Guerau-de-Arellano et al. 2005; Figure
2). In Lyme disease, two chemokines are said to stand out above the rest: CXCL2
and CCL2. CXCL2 is a neutrophil chemoattractant while CCL2 is a monocyte-macrophage
chemoattractant (Guerau-de-Arellano et al. 2005). B. burgdorferi
is believed to activate these recruited monocytes to secrete the cytokines interleukin-ß
and tumor necrosis factor-α (TNF-α) (Franz et al. 1999).
Lyme arthritis has characteristics inflammatory cytokines associated with it.
One of the predominant cytokines present in Lyme arthritis is CXCL1. This neutrophil
chemoattractant was shown to be essential to Lyme arthritis development, as
mice lacking the CXCL1 receptor developed low-severity arthritis (Guerau-de-Arellano
et al. 2005). Another cytokine found to be important in the development
of Lyme arthritis is the cytokine interleukin-1 (IL-1). Phagocytes have been
shown to induce the production of IL-1 when stimulated by the LPS on the spirochete
(Beck et al. 1989). The secretion of IL-1 is the major cause of joint
pain and destruction. IL-1 stimulates the secretion of prostaglandins and collagenase
from chondrocytes. Other chondrocyte secretions lead to the resorption of bone
by osteoclasts and the destruction of cartilage. IL-1 is also a fever inducing
cytokine which can induce the secretion of antibody and interleukin-2 (IL-2),
leading to the increased presence of lymphocytes and the antibodies IgM and
IgG in the joints (Beck et al. 1989).
Lyme neuroborreliosis is another component of Lyme disease that can occur once
the spirochete has metastasized and infected the nervous tissue. Typically,
the central nervous system is first infected, resulting in meningitis or an
inflammation of the brain and spinal cord. These lethal inflammations are caused
by unregulated production of the inflammatory cytokines interleukin-6 and TNF-α.
OspA is the lipoprotein thought to bind to toll-like receptors, as mentioned
earlier, to elicit this response in the nervous tissue by dysregulating the
mitogen activated protein kinase pathway (Ramesh et al. 2005).
Untreated or treatment-resistant Lyme disease can lead to autoimmune disease, especially in the case of Lyme arthritis. The disease is typically manifested as rheumatoid arthritis (RA) associated with the hypervariable region of the MHC Class II gene HLA-DRB1*0401 encoded within the major histocompatibility complex (Gross et al. 1998). This response occurs as a result of a cross-reactive response between OspA and self-antigen, possibly identified as LFA-1. This is a result of the secretion of the cytokine interferon-γ (IFN-γ). IFN-γ upregulates the expression of intercellular adhesion molecule 1 (ICAM1) to bind and recruit via extravasation LFA-1 expressing cells such as activated TH1 cells as well as upregulates the presentation of antigen in the context of MHC Class II by antigen presenting cells (APCs) (Gross et al. 2000). As a result of increased TH1 cells and APCs, the risk of LFA-1 presentation by APCs that have phagocytosed self-antigen increases, resulting in an immune response to this self-antigen . In this manner, even after the spirochete has been eliminated, there is still this autoreactive self-antigen that will cause an autoimmune response. (Gross et al. 1998).
Treatment
Antibiotic treatment as early as possible will help alleviate most of these
symptoms and eradicate the disease from the body. Early treatments have the
widest range of oral antibiotics indicated to treat Lyme disease: tetracycline,
amoxicillin, doxycycline, erythromycin and penicillin. Later stage Lyme disease
tends respond best, if at all, to penicillin or ceftriaxone (Figure 5). These
antibiotics are given for typically two weeks to a month, with cephalosporins
and ceftriaxone yielding the best results (Johnson et al. 1990).
The FDA approved the vaccine under the band name LYMErix in early 1999. The
major target of these vaccines is the outer surface protein OspA. A three injection
vaccine was approved that injects OspA with adjuvant in three stages, with a
49% efficacy after two injections and a 76% efficacy after three injections.
This vaccine was also shown to be effective in preventing a late, chronic infection
(Steele et al. 1998). The mechanism of action was found to be truly
novel: OspA plus adjuvant is injected into human muscle tissue where the subject
begins to develop anti-OspA IgG antibodies against the specific epitope LA-2
on OspA. When infected ticks feed on human skin, these antibodies interact with
B. burgdorferi while still in the midgut of the tick before the upregulation
of OspC occurs (Rx List 2004).
However, this vaccine does include some limitations. First, it has not been approved or thoroughly studied in individuals less than 15 years of age and is not indicated for pediatric use. Also, the stability of OspA and its rate of change is unknown and may effect is effectiveness over time. In addition, the vaccine is costly, as Lyme disease is difficult to diagnose and antibiotic treatment seems to work with high efficacy and low morbidity. The Advisory Committee for Immunization Practices also stipulates the usage for those persons between the ages of 15 and 70 and for those women who are not pregnant. The Committee also says it is only to be use for those persons at high risk or persons traveling to endemic areas during high season (Thanassi et al. 2000).

Figure 5. Antibiotic regimens for Lyme disease at various stages (Steere 2001)
Prevention
Precautions should be taken to those who live endemic areas at risk for exposure
to ticks. These include:
References.
Beck, G., Benach, J.L., Habicht, G.S. (1989). Isolation of interleukin 1 from joint fluids of patients with Lyme disease. The Journal of Rheumatology 16: 800-806.
Bubeck-Martinez, S. (2005). Immune evasion of the Lyme disease spirochetes. Frontiers in Bioscience 10: 873-878.
Franz, J.K., Priem, S., Rittig, M.G., Burmester, G.R., Krause, A. (1999). Studies on the pathogenesis and treatment of Lyme arthritis. Wiener Klinische Wochenschrift 111: 981-984.
Frey, R.J. (2002) Lyme diease. The Gale Encyclopedia of Medicine 3: 2057-2061
Friedlander, E. (2006). Infectious Diseases. <http://www.pathguy.com/lectures/infect.htm> Accessed April 23, 2006
Gross, D.M., Forsthuber, T., Tary-Lehmann, M., Etling, C. Ito, K., Nagy, Z.A., Field, J.A., Steere, A.C., Huber, B.T. (1998). Identification of LFA-1 as a candidate autoantigen in treatment-resistant Lyme arthritis. Science 281: 703-706.
Gross, D.M., Huber, B.T. (2000). Cellular and molecular aspects of Lyme arthritis. Cellular and Molecular Life Sciences 57: 1562-1569.
Guerau-de-Arellano, M., Huber, B.T. (2005). Chemokines and Toll-like receptors in Lyme disease pathogenesis. TRENDS in Molecular Medicine 111: 114-120.
Johnson, R.C., Kodner, C.B., Jurkovich, P.J., Collins, J.J. (1990). Comparative in vitro and in vivo susceptibilities of the Lyme disease spirochete Borrelia burgdorferi to cefuroxime and other antimicrobial agents. Antimicrobial Agents and Chemotherapy 34: 2133-2136.
Ranesh, G. Philipp, M.T. (2005). Pathogenesis of Lyme neuroborreliosis: mitogen-activated protein kinases Erk1, Erk2, and p38 in the response of astrocytes to Borrelia burgdorferi lipoproteins. Neuroscience Letters 384: 112-116.
Rx List. 2004 December 8. LYMErix. <http://www.rxlist.com/cgi/generic/lymerix.htm> Accessed 2006 April 23
Steele, A.C., Sikland, V.K., Meurice, F., Parenti, D.L., Fikrig, E., Schoen,
R.T., Nowakowski, J.,
Schmid, C.H., Laukamp, S., Buscarino, C., Krause, D.S. (1998). Vaccination against
Lyme disease recombinant Borrelia burgdorferi outer-surface lipoprotein
A with adjuvant. The New England Journal of Medicine 339: 209-215.
Steere, A.C. (2001). Lyme disease. The New England Journal of Medicine 345: 115-125.
Thanassi, W.T., Schoen, R.T. (2000). The Lyme disease vaccine: conception, development, and implementation. Annn Intern Med. 132: 661-668.
Yanagihara, Y., Masuzawa, T. (1997). Lyme disease (Lyme borreliosis). FEMS
Immunology and Medical Microbiology 18: 249-261.
Created by: Joseph
Scognamiglio ![]()
Return to: Immunology Homepage
Return to: Davidson College Homepage