This web page was produced as an assignment for an undergraduate course at Davidson College.
TAP
(Transporter Associated with Antigen Processing)
Background Information on TAP
The transporter associated with antigen processing, or TAP, transfers peptides from the cytosol to the lumen of the endoplasmic reticulum so that peptides can be loaded onto MHC I molecules and presented on the cell surface. TAP is an example of an ATP-binding cassette (ABC) transporter, which is the largest and most diverse family of membrane-spanning transport proteins. ABC transporters transport diverse molecules, from large proteins to ions, across plasma membranes, and are found in the animal and plant kingdom, bacteria, and archea (Buttgereit,2002).
Role of TAP in Function of Immune System
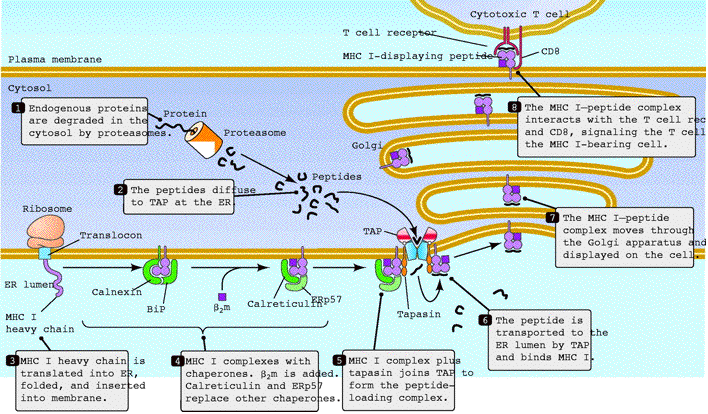
Figure 1: Overview of MHC I Antigen-Processing Pathway. TAP becomes associated with tapasin, MHC I, calreticulin, calnexin, and ERp57 in the peptide-loading complex. TAP transports cytosolic peptides into the Endoplasmic Reticulum, where peptides become associated with MHC I and eventually the MHC I-peptide complex is presented on the surface of a cell. Figure taken from Abele et al.( 2004), permission pending.
In order for cytotoxic T cells to kill cells that are infected with viruses or bacteria that live in the cytosol, MHC class I molecules must present peptides originating in the cytosol onto the cell surface to be recognized by CD8+ cytotoxic T cells. TAP functions during the process of loading cytosolic proteins onto MHC Class I described below. After a proteosome degrades endogenous proteins into a 3-22 residue peptide in the cytoplasm, a small fraction of the peptides diffuse to TAP at the ER. Meanwhile, MHC class I heavy chains assemble and bind to calnexin, a chaperone protein, in the ER membrane. Then B2-microglobulin binds to the MHC class I heavy chain and the dimer separates from calnexin and binds to calreticulin and Erp57 to form the MHC class I loading complex. Next, tapasin binds the complex to form the peptide-loading complex and join the complex with TAP. Then the degraded cytosolic peptides are transported by TAP into the lumen of the ER and loaded onto an MHC I molecule. It is uncertain whether the TAP:tapasin complex directly loads peptides onto MHC class I molecules or whether the MHC class I molecule scans the peptides once they are transported into the ER. The peptide:MHC complex leaves the endoplasmic reticulum and then traverses the Golgi apparatus to the cell surface. The MHC I-peptide complex interacts with the TCR and CD8 of a cytotoxic T cell, allowing the T-cell to kill the infected cell and carry out the immune response. In normal, uninfected cells, the MHC class I molecules bind to self proteins and go to the cell surface. When the cell becomes infected by a virus, the MHC I presents pathogen-derived peptides at the cell surface(Abele et al., 2004, Janeway et al., 2005).
Role of TAP in Peptide-Loading Complex
TAP becomes associated with tapasin, MHC I, calreticulin, calnexin, and ERp57 in the peptide-loading complex. Since the assembly of MHC I with ERp57, calreticulin,and tapasin is less efficient without TAP, TAP is a chaperone for members of the loading complex(Abele et al., 2004). The TAP-MHC I interaction in the peptide complex may help stabilize MHCclass I:Beta-2 Microglobulin (Owen et al., 1999). The peptide loading complex has more lateral mobility while TAP is inactive and less lateral mobility while TAP is translocating peptides. In addition, the nucleotide binding site of TAP may transmit a signal for the peptide-loaded MHC I to dissociate from the complex(Abele et al., 2004).
Location of TAP
Immunoelectron and immunofluorescence microscopy have localized TAP1 and TAP2 in the ER and cis-Golgi(Buttgereit et al., 2002).
Structure and Function of TAP
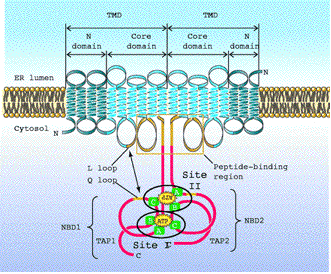
Figure 2: Schematic Model of TAP. This figure illustrates the TMD, peptide-binding region, NBD, and other important parts of TAP. Fi
gure taken from Abele et al.( 2004), permission pending.
TAP is an ATP-binding cassette (ABC) transporter, consisting of two subunits, TAP1 and TAP2, both of which contain a C-terminal ABC ATPase domain and an N-terminal transmembrane domain (TMD)(Gaudet et al., 2001). Both subunits must be present for TAP to function, and both subunits hydrolyze ATP to power peptide transport. The hydrophobic TMD consists of a core of 6 helices along with an NH2-terminal extension of 4 and 3 helices for TAP1 and TAP2 respectively (Abele, 2004). The peptide-binding region consists of the last cytosolic loop and 15-amino acids from the last transmembrane helix of TAP1 and TAP2 while the tapasin-recruiting region consists of the NH2-terminal extensions(Nijenhuis, 1996). The Walker A and B sequences, along with the C loop sequence, of the nucleotide-binding domains, are involved in binding and/or hydrolyzing ATP. The Nucleotide-Binding Domains(NBD) of TAP also convert the energy of ATP hydrolysis to peptide transport. Site I consists of the ATP binding pocket composed of residues of the Walker A and B motifs of NBD1 and the C loop of NBD2. Researchers propose that the Q and L loops interact with each other(Abele, 2004).
Although the ability of a peptide to bind to TAP does not require ATP, peptide transport requires ATP hydrolysis. TAP does not translocate all peptides with equal efficiency. It chooses peptides according to their length and their sequence, which affects the pool of peptides available for MHC I binding (Nijenhius, 1996). TAP binds peptides of 8-16 amino acids and transports peptides of 8-12 amino acids efficiently. However, TAP may transport peptides as long as 40 amino acids. The first three residues and the COOH-terminal residues are essential for binding(Abele, 2004). TAP, like MHC I, prefers peptides with hydrophobic or basic residues at the carboxy-terminus and has a bias against proline in the first three amino-terminal residues(Janeway et al., 2005). The sequence between the ends of the peptides can vary significantly, which allows optimal peptide diversity to protect an organism from a wide variety of pathogens(Abele, 2004). The following image illustrates the cycle of events that occur when TAP transports a peptide from the cytosol into the ER.
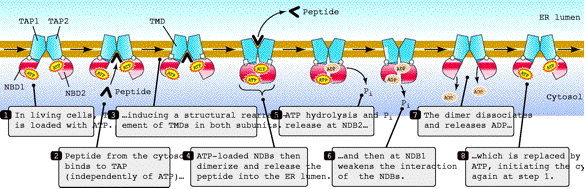
Figure 3: Proposed Peptide Transport Cycle of TAP. This figure illustrates each step involved in the transport of peptides from the cytosol into the ER by TAP. Figure taken from Abele et al.( 2004), permission pending.
Genomics of TAP
The genes for human TAP1 and TAP2 are located in the MHC II locus of chromosome 6 and they comprise 8-12 kb each. Deletions of either or both of the TAP genes result in greatly reduced surface expression of MHC class I molecules and failure to present cytosolic antigens to cytotoxic T cells. In both human and mice, TAP genes are polymorphic; therefore, several different alleles of human TAP1 and TAP2 exist(Buttgereit et al., 2002). Viral infections produce interferons which induce TAP1 and TAP2 genes to increase MHC I presentation on the surface of a cell(Janeway et al., 2005).
Mutations/Diseases in TAP
Cells with a mutation in TAP1 or TAP2 express abnormally low levels of MHC class I molecules on the surface because MHC class I molecules are unstable without a bound peptide. Several viruses interfere with TAP to decrease the presentation of MHC I, and thus, avoid an immune response. Herpes simplex virus type 1(HSV-1) encodes the protein ICP47, which binds the peptide binding site of TAP to block the first step in the translocation pathway(Tomazin et al., 1998). The critical amino acids of ICP47 inhibit the NH2-terminal region from residue 3 to 34(Buttgereit et al.,2002). This interaction between TAP and ICP47 is species specific(Tomazin et al., 1998).
Human cytomegalovirus (HCMV) encodes an ER-resident glycoprotein, gpUS6, which binds to the ER-luminal part of the TAP complex and inhibits the TAP-mediated peptide transport. gpUS6 does not affect peptide binding or the association of tapasin, calreticulin, and MHC I molecules with TAP. gpUS6 also binds to TAP and stabilizes TAP1 conformation so TAP cannot bind ATP, which inhibits peptide translocation. IFN-Gamma induction counters gpUS6’s inhibition of peptide transport, but HCMV expresses other proteins that interfere with MHC I presentation in a different manner(Buttgereit et al.,2002).
Adenoviruses encode E3/19K, a protein that binds MHC class I molecules directly and traps them in the ER. E3/19K also binds to TAP to inhibit tapasin action, which keeps TAP from associating with MHC I. Epstein-Bar Virus(EBV) avoids immune recognition by secreting vIL-10 to downregulate expression of TAP 1, which decreases MHC class I cell surface expression(Buttgereit et al.,2002).
In Type II Bare Lymphocyte Syndrome (BLS), a nonfunctional TAP2 caused by a premature stop codon results in reduced expression of MHC I on the cell surface and reduced amounts of cytotoxic T lymphocytes. However, a small fraction of antigenic peptides can be transported into the ER lumen without the TAP pathway(Buttgereit et al., 2002). Both TAP1 and TAP2 deficiencies cause chronic inflammation of respiratory tract in late childhood and vasculitis, but a person can often live asymptomatically for several decades(de La Salle et al., 1999). Eventually, recurrent bacterial infections may cause lung damage, respiratory failure, and even death. Since MHC I is ubiquitous throughout tissues, gene therapy would not work against TAP-deficient syndromes(Buttgereit et al., 2002).
TAP deficiencies may contribute to MHC-linked autoimmune diseases such as type I diabetes, Sjorgen’s syndrome, and Grave’s disease. In these diseases, improper transcription of TAP genes can decrease MHC Class I cell surface expression, and therefore, make these cells a target for natural killer cells.
Murine and human cancers exhibit a downregulation or mutation of TAP, which leads to a loss or decrease in MHC I surface expression and the ability to avoid immune recognition. This downregulation of TAP in several malignancies can be corrected with cytokines like IFN-Gamma and TNF-Alpha(Buttgereit et al., 2002).
Drugs Involving TAP
Treatment with several cytokines, such as IFN-Gamma and TNF-Alpha, can upregulate TAP, resulting in a phenotypic increase of MHC class I expression. However, the cytokine interleukin-10 can lead to TAP downregulation. A study using a vaccinia virus containing the TAP1 gene decreased tumors in mice and, therefore, showed that exogenous TAP could be used in cancer therapies(Buttgereit et al., 2002).
Works Cited
Abele, Rupert and Robert Tampe. "The ABCs of Immunology:Structure and Function of TAP, the Transporter Associated with Antigen Processing" Physiology 19 (2004): 216-224.
Buttgereit, Brigitte and Robert Tampe. "The Transporter Associated With Antigen Processing:Function and Implications in Human Diseases." Physiological Reviews 82 (2002): 187-204.
de la Salle H, et al. "HLA Class I Deficiencies Due to Mutations in Subunit 1 of the Peptide Transporter TAP1." The Journal of Clinical Investigation. 103(1999): R9-R13.
Gaudet R, Wiley D.C. "Structure of the ABC ATPase Domainof human TAP1, the Transporter Associated with Antigen Processing". The EMBO Journal. 20(2001): 4964-4972.
Janeway, C.A., Travers, P., Walport, M., Schlomchik, M. Immunobiology 6th Ed: The Immune System in Health and Disease. New York: Garland Publishing, 2005.
Nijenhuis, M. and GJ Hammerling. "Multiple Regions of the Transporter Associated with Antigen Processing (TAP) Contribute to its Peptide Binding Site." The Journal of Immunology 157 (1996): 5467-5477.
Owen, Barbara A. and Larry Pease. "TAP Associated Influences the Conformation of Nascent MHC Class I Molecules." The Journal of Immunology 162 (1999): 4677-4684.
Tomazin, Roman, et al. "Herpes Simplex Virus Type 2 ICP47 Inhibits Human TAP but not Mouse TAP" The Journal of Virology 72 (1998): 2560-2563.
If you have any questions or concerns about this web page please contact Ryan.
Return to Davidson College Biology Home Page.
Return to Immunology Home Page.
Return to Ryan's Immunology Home Page.