INTRODUCTION
Cell-cell interactions and signal transductions have become topics of great interest in cellular and molecular biology. There are a large number of biological processes ranging from fertilization and embryonic development to viral infections that are dependent upon a complex interplay between cells which ultimately leads to the fusion of their plasma membranes (Wilson and Snell, 1998). Despite this common cellular phenomenon, there is not much known about the molecular mechanisms responsible for cell-cell fusion.
The haploid unicellular biflagellated green alga, Chlamydomonas reinhardtii, is a model organism for studying the cellular and molecular mechanisms that allow cells to recognize one another, produce mating type specific structures, and ultimately fuse to form zygotes. C. reinhardtii is commonly found among soil and fresh water habitats and spends most of its life cycle as a haploid mitotic (vegetative) cell. C. reinhardtii cells obtain the ability to participate in sexual interactions during gametic differentiation, which is a developmental process stimulated by nitrogen deprivation and light. There are two separate recognition systems that regulate the mating of C. reinhardtii gametes. Hydroxyproline-rich glycoproteins known as flagellar agglutinins mediate the first system by extrinsically associating with the flagellar surface and instigating the initial contact between gametes (Goodenough, et al., 1995). The individual gametes of C. reinhardtii are of two different mating types, plus and minus, which appear to express either plus or minus traits such that a given cell will display only one type of agglutinin (Adair, 1985). Plus and minus agglutinins are homologous proteins that are of the same family as the cell wall of C. reinhardtii and of the cell-wall hydroxyproline-rich proteins of higher plants (Longo, 1987, Russell and Damas, 1992).
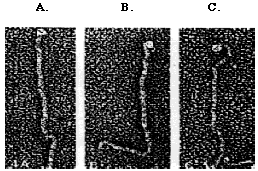
Gametic differentiation is assessed by mixing gametes of the opposite mating type together. Unlike vegetative cells, gametic cells will clump together. The gametes of C. reinhardtii mate by a process called sexual agglutination in which the flagellar agglutinins allow two gametes ofopposites mating types to adhere to one another. The specific mechanisms of mating-type recognition and gamete adhesion have yet to be completely described; however, both mating type agglutinins have been isolated (Adair, 1985). The mt+ agglutinin has a straight shape, whereas, the mt- agglutinin is hooked at one end (Figure 1).
Figure 1. The morphology of agglutinin polypeptides. mt+ (A and B), mt- (C). The mt- agglutinin has a hooked end, whereas, the mt+ agglutinins have straight ends (Micrograph by John Heuser; Adair, 1985).
When the fertilization tubule of the plus mating structure makes adhesive contact with the minus mating structure, the plasma membranes of the gametes are able to fuse and yield heterokaryotic quadriflagellated zygotes. When a zygote matures, the cell undergoes nuclear fusion, chloroplast fusion, the resorption of flagella, and the synthesis of a zygote-specific cell wall. Under the right conditions, the zygote is able to undergo meiosis and germination to yield four haploid progeny, two mt+ and two mt- (Harris, 1989) (Figure 2).
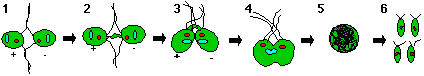
Figure 2. The mating process of C. reinhardtii. 1) mt+ and mt- cells adhere to one another via flagellar agglutinins and are signalled to release cAMP 2) cAMP messenger system signals cells to produce mating structures 3) cells attach via their mating structures 4) cells fuse 5) a zygote forms and 6) under appropriate conditions the zygote will undergo meiosis and germination to yield four haploid progeny (model adopted from Harris, 1989).
C. reinhardtii is particularly well suited for classical genetic analysis (Quarmby and Hartzell, 1994). New mutant strains and gene clones are being generated at a rapid rate by insertional mutagenesis and plasmid rescue (Tam and Lefebvre, 1993; van Winkle-Swift, 1992). Dr. Virginia Armbrust of Washington University isolated a mt- mutant strain of C. reinhardtii called GA1. Dr. Armbrust's lab has focused its research efforts on the molecular dissection of this strain of cells in order to determine why they cannot mate properly with wild type mt+ or mt- cells. When GA1 cells are mixed with wild type mt+ cells, gametic differentiation cannot be assessed because flagellar agglutination does not take place. However, when cells are treated with cAMP, cell walls are shed and zygotes are formed, indicating that the second recognition system is functional (personal communication; Pasquale and Goodenough, 1987).
GA1 was isolated when Armbrust was performing a mutagenesis screen with an auxotrophic mutant, arg-7 mt-, strain of C. reinhardtii . Because this strain of cells lacks the arginine 7 gene, they can only grow on arginine supplemented media. Armbrust transformed arg-7 mt- cells with a linearized plasmid (pArg7.8) containing the arginine 7 gene expecting that the DNA would be incorporated into the host genome. When she plated the cells onto medium lacking arginine, she isolated approximately 1000 strains of cells that successfully incorporated the gene. The transformed cells were screened for behavioral mutations. When she tried individually mating these 1000 stains of mt- cells with wild type mt+ gametes, she discovered that one strain (GA1) did not fuse with a mt+ gamete to form a zygote. The flagella of GA1 gametes are not able to adhere to the flagella of the plus gametes which lead to several questions: What prevents the mutant strain from agglutinating? Is the mt- agglutinin still being synthesized? Is the agglutinin being properly transported to the flagella? Does the phenotype have something to do with the glycosylation of agglutinin?
Because the inserted DNA often retains plasmid sequences that code for ampicillin resistance and the E. Coli origin of replication, flanking C. reinhardtii DNA can be cloned by transforming E. coli with the C. reinhardtii DNA and selecting for ampicillin resistance (Quarmby and Hartzell, 1994). This process, called plasmid rescue, was used to isolate a portion of the mutant gene from GA1 which was then used as a probe to isolate the wild type GA1 gene. Once isolated, the DNA was digested with eight different restriction enzymes (Bam HI, Eco RI, Not I, Sal I, Sac I, and Xho I) in order to create a restriction enzyme map of the gene (Figure 3).
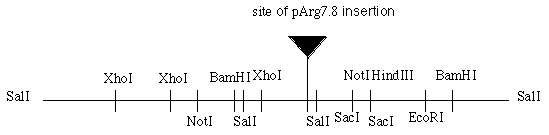
Figure 3. Restriction map of the putative GA1 gene and location of pArg 7.8 insertion. Total length is 12.1 kilobases (Dr. Virginia Armbrust, personal communication).
RNA blot hybridizations were performed using the isolated mRNA from wild type mt- cells. Five different pieces of the putative gene (a-e) were used as probes and it was discovered that the probes bound to one or two mRNAs (figure 4). For instance, the 7.0 kb Bam HI/ Bam HI fragment (probe b) revealed the presence of two mRNA bands, one of about 1.8 kb and one of about 4.4 kb. Probes a, d, and e all hybridize to the 4.4 kb message and probe c only hybridizes to the 1.8 kb message.
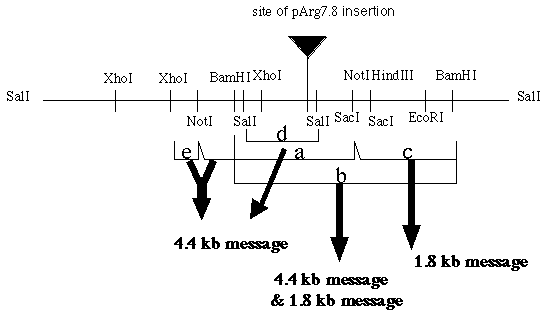
Figure 4. The five fragments (a-e) isolated from the putative gene. Each fragment was used as a probe. Each fragment hybridized to one or two mRNAs (re-drawn from Armbrust's diagram).
Because probe b was found to hybridize to both the 4.4 and 1.8 kb message, it was used to probe GA1 mRNA which revealed only one band of about 1.8 kb. The 4.4 kb message was missing from the GA1 gametes and vegetative cells but was present in all wild-type cells. These results were confirmed by using probe e on a GA1 mRNA blot analysis (Figure 5). A wild type mt- cDNA library was probed with fragment e and as a result, four cDNAs of varying lengths (900, 700, 1200, and 1900 bp) were isolated.
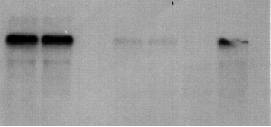
Figure 5. Image of a northern blot that used fragment e to probe wild-type plus vegetative cells (lane 1 from left), wild-type minus vegetative cells (lane 2), GA1 vegetative cells (lane 3), wild-type plus gametes (lane 4), and wild-type minus gametes (lane 5), GA1 gametes (lane 6), and 24 hour wild- type zygotes (lane 7). In lanes 1 and 2, the 4.4 kb mRNA is easily detected and is weakly detected in lanes 4 and 5. The mRNA is present in lane 7 as well. The 4.4 kb message is not detected in lanes 3 and 6 (data provided by Dr. Virginia Armbrust).
Dr. Armbrust's lab continues to isolate more cDNA and to sequence all of the cDNA that has already been isolated. The current study was designed to continue the molecular characterization of GA1 and in effect continue the molecular dissection of cell-cell interactions of C. reinhardtii. In the process of sequencing the cDNA, the sequence appeared to be similar to an H+-ATPase which lead us to believe that GAI was involved in maintaining low pH levels in endosomal compartments and as a result, involved in facilitating the glycosylation process. Chloroquine, which raises intracellular pH, and folimycin, an H+-ATPase inhibitor, were individually used on wild type cells to try to chemically mimic the GA1 phenotype. Our data from these experiments and further cDNA sequencing indicate that these drugs do not mimic the GA1 phenotype and that GA1 is more similar to a plasma membrane H+-ATPase. We have subcloned half of the putative GA1 gene and Dr. Armbrust is in the process of subcloning the other half for transformation experiments to complement the GA1 mutation. We were also interested in using tetrad analysis to determine if GA1 is linked to the sex chromosome (VI) and if mt- cells are the only cells that can have the GA1 phenotype.