Human Molecular Genetics, Imperial Cancer Research Fund,
Lincoin's Inn Fields, London WC2A 3PX, UK
t Laboratoire d'lmmunogenetique
Humaine, INSERM U. 276,
Institut Pasteur, 25/28 rue du Dr Roux, 75724 Paris Cedex 15,
France
THE testis-determining factor gene (TDF) lies on the Y chromosome and is responsible for initiating male sex determination. SRY is a gene located in the sex-determining region of the human and mouse Y chromosomes and has many of the properties expected for TDF1-3. Sex reversal in XY females results from the failure of the testis determination or differentiation pathways. Some XY females, with gonadal dysgenesis, have lost the sex-determining region from the Y chromosome by terminal exchange between the sex chromosomes4 or by other deletions5. If SRY is-TDF, it would be predicted that some sex-reversed XY females, without Y chromosome deletions, will have suffered mutations in SRY. We have tested human XY females and normal XY males for alterations in SRY using the single-strand conformation polymorphism assay6,7 and subsequent DNA sequencing. A de novo mutation was found in the SRY gene of one XY female: this mutation was not present in the patient's normal father and brother. A second variant was found in the SRY gene of another XY female, but in this case the normal father shared the same alteration. The variant in the second case may be fortuitously associated with, or predisposing towards sex reversal; the de novo mutation associated with sex reversal provides compelling evidence that SRY is required for male sex determination.
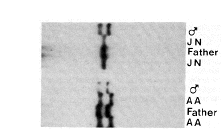
SRY sequences were amplified by the polymerase chain reaction from DNA of XY females, as well as normal male controls. The amplified products were cleaved with restriction enzymes and run on non-denaturing polyacrylamide gels to detect single-strand conformational polymorphisms (SSCP) (Fig. 1). Three patterns were detected: one pattern common to all of 50 normal male controls and the majority of patient samples; a second pattern for XY female individual (A.A.) and a third pattern for XY female (J.N.). The normal father and brother of A.A. were tested and found to give the same pattern as the controls, implying that A.A. has suffered a de novo mutation. In contrast DNA from father (N.) and his XY daughter (J.N.) showed the same pattern. The amplified products (609 base pairs) from both families were cloned and sequenced (Fig. 2a). As predicted by the SSCP assay, A.A. has a de novo mutation G->A that is not shared with her father and brother. For J.N., a G-> C change was shared with her father. Other than these observed base changes, the XY females J.N. and A.A. had complete sequence identity to SRY over the amplified 609 base pairs (bp) (Fig. 2a). Paternity in both families was confirmed by Southern blotting with minisatellite probes8,9.
The de novo mutation in A.A. causes a conservative change from methionine to isoleucine at a residue that lies within the putative DNA-binding motif of SRY and is identical in all SRY and SRY-related genes (Fig. 2b). The association of a de novo mutation with a new phenotype provides evidence that the phenotype and mutation are related, and by inference that SRY is required for male sex determination. Formally this does not exclude the possibility that other genes on the Y chromosome are required for sex determination, but previous work suggests that, if they exist, these genes must be located adjacent to the pseudoautosomal boundary, close to SRY1,10.
The variant found in J.N. causes a conservative change from valine to leucine (Fig. 2b). This residue is conserved amongst SRY and SRY-related sequences with the exception of a mouse autosomal gene (autosomal-4), which has a conservative change to isoleucine at this position2. There are several possible explanations for this variant. First, the variant could cause conditional sex reversal depending on other genetic or environmental factors. A similar observation has been described in the mouse, where the ability of some alleles of Tdy to induce testis formation depends on the genetic background11. Second, the variant could be fortuitously found in a family segregating for an autosomal or X-linked sex reversing gene. Finally, the variant could cause sex reversal and the father is mosaic for wild-type and variant sequences.
The SRY gene of the majority of the XY females we have tested appears normal by the SSCP assay. It is possible that these individuals have mutations in SRY that are not detected by the assay we have used, either because they do not cause a band shift or because they fall outside the region tested. Alternatively, these individuals may have mutations in another part of the sex-determining pathway.
FIG. 1 SSCP analysis of SRY in XY females. On the left is family
N: ![]() , DNA from the normal 46XY male
cell line PGF12; DNA from the
father is flanked on each side by DNA of his XY daughter (J.N.).
On the right is family A: again DNA from the father is flanked
by that of his XY daughter (A.A.). In total, 11 XY females were
tested by SSCP. These individuals: P.F., J.A., A.M., R.B., GM2598,
A.S., M., A.A., I.D., J.N. and K.L. all have gonadal dysgenesis,
are karyotypically normal and positive both for the Y-pseudoautosomal
boundary and ZFY . A.A. and J.N. both have pure gonadal
dysgenesis: A.A. has a streak gonad with ovarian stroma and no
germ cells; J.N. has a cystic gonad with ovarian stroma and no
germ cells. The fathers, A. and N. are both normal, fertile males.
Fifty normal males were tested as controls and no variation detected.
, DNA from the normal 46XY male
cell line PGF12; DNA from the
father is flanked on each side by DNA of his XY daughter (J.N.).
On the right is family A: again DNA from the father is flanked
by that of his XY daughter (A.A.). In total, 11 XY females were
tested by SSCP. These individuals: P.F., J.A., A.M., R.B., GM2598,
A.S., M., A.A., I.D., J.N. and K.L. all have gonadal dysgenesis,
are karyotypically normal and positive both for the Y-pseudoautosomal
boundary and ZFY . A.A. and J.N. both have pure gonadal
dysgenesis: A.A. has a streak gonad with ovarian stroma and no
germ cells; J.N. has a cystic gonad with ovarian stroma and no
germ cells. The fathers, A. and N. are both normal, fertile males.
Fifty normal males were tested as controls and no variation detected.
METHODS. Polymerase chain reactions were performed with the primers
XES7 and XES2 located within the SRY open reading frame, amplifying
a 609-bp fragment. The primer sequences are: XES7, 5'CCCGAATTCGACAATGCAATCATATGCTTCTGC3';
XES2, 5'CTGTAGCGGTCCCGTTGCTGCGGTG3'. PCRs were performed with
~100 ng of genomic DNA, 200 µM each dNTP, 0.5 µM each
primer, 1.5 mM MgCl2, 10 mM Tris (pH 8.3), 50 mM KCl,
0.01% (w/v) gelatin, 0.25 U of Taq polymerase and 0.5 µl
of [a-32P]dCTP (3,000 Ci mmol-1,
10 mCi ml-1) in a volume of
10 µl. Reactions were cycled for 1.2 min at 94° C, 1.2
min at 65° C and 2 min at 72° C for 35 cycles. One µl
of the product was digested with HinfI and TaqI in the presence
of 4 mM spermidine hydrochloride in a 10 µl volume. The
digested DNA was diluted 1:10 in 0.1% SDS, 10 mM EDTA, followed
by a 1:2 dilution in 95% formamide, 20 mM EDTA, 0.05% bromophenol
blue, 0.05% xylene cyanol. Samples of 1-3 µl were heated
at 80° C for 5 min to denature the DNA, then loaded onto 6%
acrylamide, 10% glycerol non-denaturing gels using a sequencing
gel apparatus. Electrophoresis was carried out at 25 mA, with
a fan heater set on cold directed at the gel as a cooling device.
Autoradiography of the dried gels was for 3 days without an intensifying
screen.
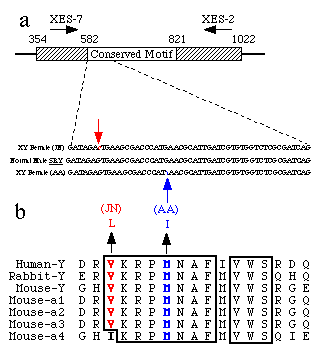
FIG. 2 a, The open reading frame of the genomic clone pY53.3
(SRY) is shown extending from 354-1,022 bpt. The conserved motif,
which encodes a potential DNA-binding protein, extends from 582
bp to 821 bp1. The primers XES-7
and XES-2 were used to amplify a 609-bp region. The entire amplification
product was sequenced, but only the small region containing the
changes in the XY females is displayed. The dotted line indicates
the location within the open reading frame of the nucleotide sequence
shown. Top line, nucleotide sequence for XY female (J.N.) with
base changes from G -> C indicated by arrow. Middie line, normal
male SRY nucleotide sequence. Bottom line, XY female (A.A.) with
a base change from G -> A indicated by arrow. b, The amino-acid
sequence of SRY for the human-Y, rabbit-Y, mouse-Y and mouse autosomal
SRY-like genes a1, a2, a3,
a412.
Boxed shaded regions show identical amino acids conserved across
these species. In the XY female J.N. the variant causes a change
from valine to leucine, whereas in the XY female A.A. the mutation
causes a change from methionine to isoleucine.
METHODS. PCR products were subcloned in pUC18 vectors (NEB). Six
independent subclones from individuals (A.A.) and (J.N.) were
sequenced on both strands. Double-stranded DNA was sequenced by
the dideoxy chain termination method13
using synthetic oligonucleotide primers and Sequenase (USB).
Received 28 September; accepted 18 October, 1990.
*Present address: Centre de Recherehes de Biochlmie Macromoleculaire,
CNRS LP8402/lNSERM
U.249, Route de Mende, 34033 Montpellier Cedex, Franre.
¥To whom correspondence
should be addressed.
REFERENCES
1. Sinclair, A. H. et al. Nature 346, 240-244 (1990).
2. Gubbay. J. et al. Nature 346, 245-250 (1990).
3. Koopman, P., Munsterberg, A., Capel, B., Vlvian, N. & Lovell-Badge, R. Nature 348, 350-452 (1990).
4. Levilliers, K., Quæk, B., Weissenbach, J. & Petit C. Proc. Natn. Acad. Sci. USA. 88, 2296-2300 (1989).
5. Page, D. C., Fisher, E. M. C., McGillivary, B. & Brown, L. G. Nature 346, 279-281(1990).
6. Orita, M., Suzuki, Y., Sekiya, T. & Hayashi, K. Genomics 6, 874-879 (l989).
7. Orita, M., Iwahana, H., Kanazawa, H., Hayashi, K. & Sekiya, T. Proc. Natn. Acad. Sci. USA. 86 2766-2770 (l989).
8. Wong, Z., Wilson, V., Patel, I., Povey, S. & Jeffreys. A. J. Ann. hum. Genet. 51, 269-288 (1987).
9. Wong Z., Wilson, V., Jeffreys, A. J. & Thein, S. L. Nueleic Acids Res. 14, 4605-4616 (1986).
10, Palmer, M. S. et al. Nature 342, 937-939 (1989).
11. Eicher, F. M. & Washburn, L. L. A. Rev. Genet. 20, 327-360 (1986).
12. Goodfellow, P. J. et al. Ann hum Genet. 53,15-22 (1989).
13. Sanger, F., Nicklen, S. & Couison, A. R. Proc. Natn. Acad. Sci. USA.. 74, 5463-5467 (1977).
ACKNOWLEDGMENTS. We thank A. bffreys for the minisatellite probes, J. P. Raymond, A. Moss, M. Poissonnier for access to the A and N families: N. Abbas and C. Cotinot for technical assistance: J. Weissenbach and R. Lovell-Badge for useful comments and C. Middlemiss for typing the manuscript.