Science. Vol. 267 24 March 1995
RESEARCH ARTICLE
Click Here to See Cover Photo
Georg Halder,* Patrick Callaerts,* Walter J. Gehring^
Biozentrum, University of Basel, Klingelbergstrasse 70, CH-4056 Basel, Switzerland.
*The first two authors contributed equally
to this work.
^ To whom correspondence should be addressed.
The Drosophila gene eyeless (ey ) encodes a transcription factor with both a paired domain and a homeodomain. It is homologous to the mouse Small eye (Pax-6 ) gene and to the Aniridia gene in humans. These genes share extensive sequence identity, the position of three intron splice sites is conserved, and these genes are expressed similarly in the developing nervous system and in the eye during morphogenesis. Loss-of-function mutations in both the insect and in the mammalian genes have been shown to lead to a reduction or absence of eye structures, which suggests that ey functions in eye morphogenesis. By targeted expression of the ey complementary DNA in various imaginal disc primordia of Drosophila, ectopic eye structures were induced on the wings, the legs, and on the antennae. The ectopic eyes appeared morphologically normal and consisted of groups of fully differentiated ommatidia with a complete set of photoreceptor cells. These results support the proposition that ey is the master control gene for eye morphogenesis. Because homologous genes are present in vertebrates, ascidians, insects, cephalopods, and nemerteans, ey may function as a master control gene throughout the metazoa.
The eyeless (ey) mutation of Drosophila was first described in 1915 (1) on the basis of its characteristic phenotype, the partial or complete absence of the compound eyes. The ey alleles available today are recessive hypomorphs (weak alleles) and they lead to the reduction or complete absence of the compound eyes but do not affect the ocelli (simple eyes) on the head of the fly. Apparent null alleles that are lethal when homozygous have also been isolated (2), but they have been lost, and a detailed analysis of their phenotype is not available. Cloning and sequencing of the ey gene (3) have shown that it encodes a transcription factor that contains both a paired domain and a homeodomain. The ey gene is homologous to Small eye (Sey = Pax-6 ) in the mouse and to Aniridia in humans. The proteins encoded by these genes share 94 percent sequence identity in the paired domain, and 90 percent identity in the homeodomain and they contain additional similarities in the flanking sequences. Furthermore, two out of three splice sites in the paired box and one out of two splice sites in the homeobox are conserved between the Drosophila and the mammalian genes, which indicates that these genes are orthologous.
Both the mouse and the Drosophila gene have similar expression patterns during development. In the mouse, the expression of Sey is observed in the spinal cord, in discrete regions of the brain, and in the developing eye. The Sey gene is expressed from the earliest stages until the end of eye morphogenesis: first, in the optic sulcus, and subsequently in the eye vesicle, in the lens, in the differentiating retina, and finally in the cornea (4). In Drosophila, ey is first expressed in the embryonic ventral nerve cord and in defined regions of the brain. Later in embryogenesis, ey is transcribed in the embryonic primordia of the eye as soon as these cells can be detected. In subsequent larval stages, it continues to be expressed in the developing eye imaginal discs. During the third larval stage, ey expression becomes largely restricted to the part of the eye disc that is anterior to the morphogenetic furrow. This region consists of undifferentiated cells whereas posterior to the furrow the differentiating ommatidia are apparent (5). Because mutations in the mouse and Drosophila genes lead to a reduction or complete absence of all eye structures, and because these genes are similar in DNA sequence and in expression pattern even at the earliest stage of eye development, it has been suggested that ey and Sey may be the master control genes involved in eye morphogenesis (3). Furthermore, mutations in four other Drosophila genes with similar phenotypes (eyes absent , sine oculis , eye gone , and eyelisch ) do not affect the expression pattern of ey, which indicates that ey acts upstream of these other genes (6). These results are consistent with its possible role as a gene that controls eye morphogenesis, even though it may have additional functions in the developing nervous system. The cloning of the homologous genes from ascidians, cephalopods, and nemerteans (ribbon worms) suggests that this gene may be present in all metazoa (3).
Master control genes that act as developmental switches can be detected on the basis of their mutant phenotypes. Thus, homeotic mutations have identified master control genes that specify the body plan along the antero-posterior axis. These genes, which are characterized by a homeobox, are clustered in the Antennapedia (Antp ) and Bithorax Complexes in Drosophila, and in the Hox gene clusters of the mouse (7). Loss- and gain-of-function mutations in these genes lead to opposite homeotic transformations. For example, in Antp , recessive loss-of-function mutations are lethal at the embryonic or larval stage and lead to a transformation of the second thoracic segment (T2) toward the first thoracic segment (T2 --> T1). Dominant gain-of-function mutations lead to a transformation in the opposite direction, that is from the anterior head and T1 segments toward T2 (H,T1 --> T2) (8). These transformations can be explained by the combinatorial interaction of several homeotic genes in order to specify a given body segment. These genes have partially overlapping expression domains in several body segments and each segment is specified by a combination of homeobox genes, that is by a Hox code (9). By ubiquitous (ectopic) expression of Antp under the control of a heatshock promoter, we have changed the body plan of Drosophila and induced the formation of middle legs in place of the antennae, and also transformed the dorsal head capsule into structures of the second thoracic segment (H --> T2). This phenotype is similar to that observed in dominant gain-of-function mutations (10). However, it proved to be difficult to transform the more posterior body segments toward T2. Data for several homeotic genes indicate that there is competition between the ectopically expressed gene and the genes normally expressed in a given segment (11). This competition frequently leads to epistasis of the posterior over the anterior genes, and to segmental transformations that are confined to the anterior body segments.
The ey gene, which also contains a homeobox in addition to a paired box, differs from Antp and the other antero-posterior homeotic genes in that the hypomorphic loss-of-function mutation leads to a loss of the corresponding eye structures rather than to their homeotic transformation. This phenotype does not necessarily imply that ey acts as a developmental switch; it only shows that ey function is required for eye development. If, however, ey is the master control gene for eye morphogenesis, the ectopic expression of ey should induce the formation of ectopic eye structures in other parts of the body similar to the transformations obtained for Antp (10) and the other homeotic genes (11). Therefore we used the GAL4 system (12) and a heat-inducible expression vector in order to express the ey gene ectopically.
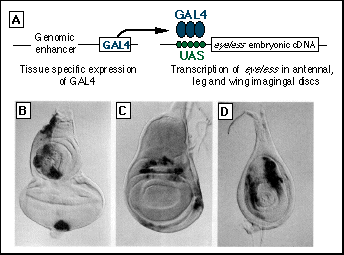
Fig. 1. Targeted expression
of ey.
(A) Schematic representation of the ectopic induction of ey by means of the GAL4
system. In (B) through (D), ß-galactosidase staining of third instar
imaginal discs (28) shows the activation of a UAS-LacZ reporter construct by the GAL4 enhancer-trap
line E132.
(B) Eye-antenna disc. The antenna portion of the disc is on the top and
the eye portion is on the bottom. ß-Galactosidase activity is detected
in parts of the antenna disc corresponding to several antenna segments and
in the periphery of the disc, which will give rise to head cuticle. The
staining observed at the most posterior part of the eye disc derives from
the optic nerve.
(C) Wing imaginal disc. ß-Galactosidase activity is detected in proximal
regions of the future wing blade, and in portions corresponding to the hinge
regions and ventral pleura.
(D) Leg imaginal disc with LacZ
expression in portions that correspond to the tibia
and femur.
Induction of ectopic eye structures. We used the GAL4 system (12) to target ey expression to various imaginal discs other than the eye discs in which ey is normally expressed. GAL4 is a yeast transcriptional activator that can activate transcription of any gene after introduction into Drosophila if the gene is preceded by a GAL4 upstream activating sequence (UAS) that consists of five optimized GAL4 binding sites ( 12). The GAL4 system is now widely used in conjunction with a method called enhancer detection (13), in which a reporter gene is provided with a weak promoter only and inserted at random sites in the genome by transposition. If the detector has inserted close to an enhancer, the reporter gene is expressed differentially. By isolating a large number of enhancer detection lines, a spectrum of different enhancers with specific temporal and spatial pattems of control can be identified. If GAL4 is used as a reporter gene, these enhancer detection lines can be used for targeted gene expression; the enhancer drives the specific expression of GAL4, which in tum can transactivate a target gene, in our case ey provided with a UAS. As indicated in Fig. 1A, the GAL4 enhancer detection line was crossed to a UAS-ey stock to generate transheterozygous flies that express ey in those cells that express GAL4. We chose approximately 20 GAL4 lines, of which only 3 gave viable adult flies to analyze in more detail (14). The results are illustrated for the GAL4 line E132. When E132 is crossed with a stock containing a UAS-LacZ construct, ß-galactosidase staining reveals the activation of the lac-Z reporter gene by GAL4 and thus the expression pattern of GAL4 in the imaginal discs. E132 expresses GAL4 in discrete regions of the wing and haltere discs, all three pairs of leg discs, and in the antennal imaginal discs (Fig. 1, B through D), which are the primordia for the respective adult structures. When the GAL4 expressing line E132 is crossed with a stock carrying an ey embryonic complementary DNA (cDNA) (15) under a GAL4-UAS control element, transheterozygous flies can be generated, and the expression of ey can be targeted into the imaginal discs as mentioned above (Fig. 1, B. C, and D for LacZ ). In the wild-type controls, ey is only expressed in the eye discs.
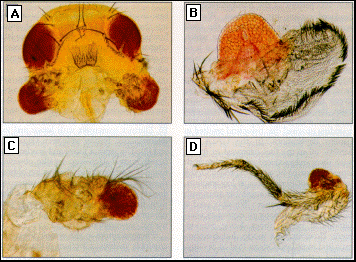
Fig. 2. GAL4 driven ectopic
expression of ey induces the formation of eye structures in various tissues. The
sites at which ectopic eyes form correspond to the regions in the imaginal
discs, in which GAL4 is expressed as assayed by the activation of a LacZ reporter construct
(Fig. 1, B. C, and D). The ectopic eye structures show ommatidial arrays,
interommatidial bristles, and red pigmentation (29).
(A) Cuticle of an adult head in which both antennae formed eye structures.
(B) Dissected wing with a large outgrowth of eye tissue. The ectopic eye
contains about 350 facets. Many interommatidial bristles are also apparent.
The normal eye contains approximately 800 ommatidia. The wing is reduced
in size. The anterior margin with its characteristic triple row of bristles
occupies most of the circumference, whereas the more posterior structures
are absent and replaced by eye tissue. The characteristic venation pattern
of the wing is disturbed by the formation of the ectopic eye structures.
(C) Dissected antenna in which most of the third antenna segment is replaced
by eye structures.
(D) Dissected middle leg with an eye-outgrowth on the base of the tibia.
As a consequence of ectopic ey expression in line E132, ectopic eye structures were induced in the wings (Fig. 2A), all six legs (Fig. 2B, for mesothoracic legs), the antennae (Fig. 2C), and the halteres. When the flies were raised at 25°C, at which temperature the cold-sensitive GAL4 is properly active, 100 percent of the transheterozygotes produced ectopic eye structures. We observed that the eye structures in the adult cuticles bulged out of the tissue in which they were induced. This phenomenon is illustrated for the wing in scanning electron micrographs (Fig. 3, B and D), and could represent sorting out of heterotypic cells in order to minimize the contact surface between the two tissue types (16). In some cases, the development of the ectopic eyes interfered with pattern formation in the surrounding imaginal disc tissue and resulted in pattern duplications. In the GAL4 line MS941, all of the flies expressed ey in the wing discs and produced eye facets on both wings. In line p339, which expressed GAL4 in a small spot in the wings in low amounts, only red pigment was formed, but again with 100 percent penetrance. We also used a heat-inducible promotor to express ey ubiquitously at various times during development. However, heat shocks during embryonic and most larval stages lead to developmental arrest. To circumvent this lethality, heat-shocks were applied after 80 hours during the middle of the third larval stage. Ectopic eye structures including complete ommatidia were induced. However, targeted ey expression by the GAL4 system was more effective.

Fig. 3. Scanning electron of
ectopic eyes (30).
(A) Scanning electron micrograph of an ectopic eye (arrowhead) in the head
region formed by the antennal disc.
(B) Overview of a fly with an ectopic eye under the wing (arrow) and on
the antenna (arrowhead).
(C) Higher magnification of (A). The ectopic eye (to the left) contains
hexagonal ommatidia and interommatidial bristles. The organization of the
facets in the ectopic eye is very similar to the pattern in the normal eye
(to the right). Some facets, however, are fused and some irregularities
in the fom of the facets are observed.
(D) Higher magnification of the ectopic eye under the wing shown in (B)
(arrow). The ectopic eye protrudes out of the thoracic body wall (ventral
pleura). The organization of the facets and interommatidial bristles are
similar to that of the ectopic eye shown in (C).
The fine structure of the ectopic eyes was analyzed by scanning electron microscopy. Well-developed ectopic eyes were most frequently observed on structures derived from the antennal and wing discs (Fig. 3, A and B). Distinct ommatidia with lenses and interommatidial bristles were seen (Fig. 3, C and D). The array of facets and bristles were largely normal. However, we also observed fusion of facets and irregular spacing of bristles in some cases. The eye structures induced on the legs were on average smaller than the ones on antennae or wings but nevertheless appeared to have a relatively normal organization.
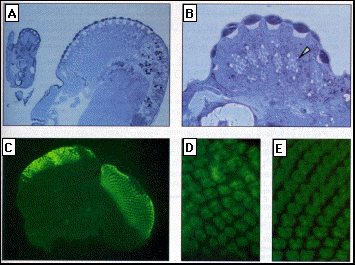
Fig. 4. Histological structure
and differentiation of photoreceptors in the ectopic eye.
(A) Micrograph of a section through an ectopic eye in the antenna (to the
left) and the normal eye (to the right) stained with Azur II and methylene
blue (15).
(B) Phase contrast micrograph of a section through an ectopic eye on the
antenna. The nomlal number and trapezoidal arrangement of the rhabdomeres
of photoreceptors is observed in the different ommatidia (arrowhead).
(C) Micrograph of an eye-antenna disc stained with an antibody against the
neuronal marker ELAV and a secondary fluorescein labeled antibody. In the
normal eye portion (to the nght), regularly spaced ommatidial clusters of
differentiating photoreceptors are detected. In the antenna part of the
disc (on the left), extensive cell proliferation has led to a doubling in
size. In this portion, a large domain of ectopically induced photoreceptors
is seen.
(D) and (E) are higher magnifications of (C), which shows the photoreceptor
clusters in the ectopic eye (D) and in the nommal eye (E), respectively.
An essentially nommal cluster fommation and cluster array is observed in
the ectopic eye.
Photoreceptors in the ectopic eyes. Microscopic analysis of sections of ectopic eye structures indicated that the ectopic ommatidia consisted of the full complement of the different types of cells and structures ( 17). In a longitudinal section of an antennal ectopic eye, we were able to distinguish cornea, pseudocone, cone cells, primary, secondary, and tertiary pigment cells, and photoreceptors with rhabdomeres (Fig. 4, A and B). At the base of the ommatidia, we observed the feet of secondary and tertiary pigment cells and a basal lamina that formed a structure with features characteristic of the fenestrated membrane of the retina. On a transverse section, the normal trapezoidal array of rhabdomeres was clearly visible (Fig. 4B, arrowhead).
We also analyzed the neuronal differentiation of photoreceptors by means of ELAV antibodies (18). Clusters of photoreceptor cells were clearly detected at ectopic sites in the imaginal discs (Fig. 4, C and D), and the sequence of neuronal differentiation observed in the normal eye disc was retained in the ectopic eye cells. A number of single cells that expressed the neuronal marker were seen at one side of an ectopic photoreceptor cluster. This expression most likely borresponds to the formation of R8 photoreceptor cells. Subsequently, groups of three, five, seven, and eight cells were detected that expressed the ELAV epitope. This series of events probably corresponds to what is observed in a normal eye disc upon passage ef the morphogenetic furrow. Thus, these observations suggest that morphogenesis of the ectopic eyes is normal and that it probably involves the formation of an ectopic morphogenetic furrow. In summary, the data presented above show that ey can induce the formation of complete and morphologically nomal ectopic eyes. It is unknown whether these ectopic eyes are functional, and whether the axons of the photoreceptors innervate the correct domains of the brain, that is, the lamina and the dorsal deuterocerebrum, respectively (19). Initial evidence suggests that the photoreceptors in the ectopic eyes are electrically active upon illumination (20).
Role of eyeless in eye morphogenesis. The reported findings indicate that ey is the master control gene for eye morphogenesis, because it can induce ectopic eye structures in at least the imaginal discs of the head and thoracic segments. The expression of ey switches on the eye developmental pathway that involves several thousand genes. The number of genes required for eye morphogenesis can roughly be estimated on the basis of the frequency of enhancer detection lines that show reporter gene expression in the eye imaginal discs posterior to the morphogenetic furrow during eye differentiation. Because approximately 15 percent of a large sample of enhancer detector lines fall into this category (21), and assuming that the Drosophila genome contains at least 17,000 genes (22), we estimate that more than 2500 genes are involved in eye morphogenesis. Our results suggest that all of these genes are under the direct or indirect control of ey, which is at the top of the regulatory cascade or hierarchy. The ey gene is expressed first and controls a set of subordinate regulatory genes, including sine oculis, another homeobox-containing gene (23). Subsequently, genes that influence cell-cell interactions and signal transduction must be regulated and, finally, the structural genes like rhodopsin, crystallin, and transducin must be expressed. The lower part of this cascade, including signal transduction pathways, has been elucidated to a large extent (24), but the upper part, and which of these interactions are direct, remain to be determined. However, ey may not only control the initial steps of eye morphogenesis, but also, as suggested from the expression pattern, it may control later steps. Thus, the same transcriptional regulator may be used at consecutive steps of morphogenesis. This could be the consequence of the conservative mode of evolution whereby the same master control gene is used repeatedly to integrate new target genes into the eye developmental pathway. In addition to eye morphogenesis, ey controls other functions in the developing nervous system, because null mutations are lethal, and the loss of eye structures alone is not the cause of lethality.
The induction of ectopic eyes in Drosophila is reminiscent of the classical experiments of Spemann (25) in which he induced ectopic eyes by transplanting the primordia of the optic cup to ectopic sites in amphibian embryos. Our experiments extend these observations and identify the gene that is necessary and sufficient to induce ectopic eyes at least in imaginal discs. In the mouse, Sey is expressed at each step of the induction process; first in the optic cup, then in the lens, and finally in the cornea, which implies that Sey may be the master control gene in the mouse eye induction process (4)
The transformation of antennal, leg, and wing tissue into eye structures by ey induction indicates that ey is a homeotic gene. In contrast to the classic homeotic genes of the Antennapedia and Bithorax Complexes, hypomorphic loss-of-function mutations in ey do not lead to homeotic transformation, but rather, they result in the loss of eye structures. However, targeted ectopic ey expression induces homeotic transformations similar to those observed in gain-of-function mutations of classic homeotic genes, like Antp . Therefore, ey represents a class of homeotic master control genes different from Antp . Gain-of-function mutants with phenotypes corresponding to those obtained in our targeted gene expression experiments have not been discovered previously.
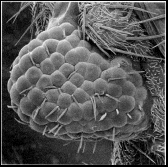
Fig. 5. The ectopic expression of mouse Pax-6 cDNA under the control of GAL4 induces the formation of ectopic eyes (26). The scanning electron micrograph shows a close-up of induced eye facets on a leg. Ommatidial arrays and interommatidial bristles very similar to the ectopic eye structures induced by the Drosophila gene (Fig. 3) were formed (30). In both cases the same GAL4 line E132 was used.
The high degree of sequence conservation between the human, the mouse, and the Drosophila genes, the similarity of the phenotypes of Aniridia, Sey, and ey, and the similarity of the expression patterns suggested to us that ey might be a master control gene for eye molphogenesis that is shared by vertebrates and invertebrates (3). Because we also found homologous genes in ascidians, cephalopods, and nemerteans we propose that ey function is universal among metazoa. In order to test whether the mouse gene can substitute for the Drosophila gene, we also used the mouse Sey gene for targeted expression in Drosophila. Similar to the results obtained for the Drosophila ey gene, the mouse gene Sey can also induce the formation of ectopic eye structures (Fig. 5) (26). As expected, the ectopic eye structures formed contain Drosophila-type ommatidia and not mouse eye structures.
Previously, the function of other mouse homeobox genes has been demonstrated in Drosophila with the use of heat inducible vectors (27). In the case of HoxB6, Drosophila legs were induced in place of the antennae (27). Obviously, the responses, but not the transcriptional regulator, are species-specific.
The observation that mammals and insects, which have evolved separately for more than 500 million years, share the same master control gene for eye morphogenesis indicates that the genetic control mechanisms of development are much more universal than anticipated. It will he informative to compare the regulatory cascade required to form a Drosophila compound eye with that of a mouse eye, to find out what the differences are, and to determine how many new genes have been recruited into these developmental pathways in the course of evolution.
REFERENCES AND NOTES
1. M. A. Hoge, Ann. Naturalist 49, 47 (1915); C. B. Bridges, Z. Biol. 4, 401 (1935).
2. B. Hochmann, Genetics 62,235 (1971).
3. R. Quiring et al., Science 265, 785 (1994).
4. C. Walter and P. Gruss, Development 113, 1435 (1991); R. Hill et al., Nature 354,522 (1991); C. C. T. Ton et al., Ceil 67, 1 059 (1 991 ).
5. R. wow and D. F. Ready, in The Deveb,ornent of Drosophila mebnogaster, M. Bate and A. MartinezArias, Eds. (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY,1993), chap.22, p.1277.
6. The expression pattern of ey was analyzed by staining of the different mutant eye-antenna discs with a polyclonal rat ey antibody and a fluorescein-bbeled secondary antibody. The mutants have been described in D. L. Lindsley and G. G. amm, The Genorne of Drosophila melanogaster (Academic Press, New York,1992), p.1133.
7. W. McGinnis and R. Krumbuf, Cell 68,283 (1992); J. Botas, Curr. Bid. 5, 1015 (1993); W. J. Gehring, M. Affolter, T. Burglin, Anna. Rev. Biochan. 63, 487 (1994).
8. W. J. Gehring, Science 236,1245 (1987).
9. E. B. Lewis, Nature 276, 565 (1978); M. Kessel, R. Balling, P. Gruss, Cell 61,301 (1990); M. Kessel and P. Gruss, ibid. 67, 89 (1991); P. Hunt et al., Nature 353, 861 (1991); P. Hunt and R. Krumlauf, Annu. Rev. Cell Biol. 8, 227 (1992).
10. S. Schneuwly, R. Klemenz, W. J. Gehring, Nature 325, 816 (1987); G. Gibson and W. J. Gehring, Devebpment 102, 657 (1988).
11. M. A. Kuziora and W. McGinnis, Cell 55,477 (1988); R. S. Mann and D. S. Hogness, ibid. 60,597 (1990); G. Gibson et al, ibid 62,1087 (1990); A. GonzalesReyes and G. Morata, ibid. 61, 515 (1990); A. Gonzales-Reyes et al., Nature 344, 78 (1990); M. Lamka et al., Development 116, 8412 (1992); J. Castelli-Gair et al. , ibid. 120,1983 (1994).
12. A. H. Brand and N. Perrimon, Devebpment 118,401 (1993); J. A. Fischer, E. Giniger, T. Maniatis, M. Ptashne, Nature 332, 853 (1988).
13. C. J. O'Kane and W. J. Gehring, Froc. NaM Acad Sci. US.A. 84, 9123 (1987); H. Bellen et al. , Genes Dev. 3, 1288 (1989); C. Wilson et al., ibid. 3, 1301 (1989); E. Bier et al. , ibid., p. 1273.
14. GAL4-crosses were established at 25°C on standard medium. The GAL41ines were crossed to a number of UAS-ey lines and most of these crosses led to embryonic or espy brval lethality. This outcome is probably the result of the ectopic expression of ey in various tissues dunng embrpogenesis because the GAL4 lines start to express GAL4 during embryonic stages. In crosses with the MS941, p339, and E132 lines, transheterozygote adults were recovered. For MS941, air ost no lethality was observed, whereas for p339 and E132, a substantial number of dead embryos or larvae were noted. For line E132, virtually only females were obtained because most males died dunng the early phases of devebpment. This result may be explained by the dependence of the bthality on the bvd of transadivation of ey by GAL4. In the E132 line, the enhancer detector construct is inserted into the X chromosome and therefore, is dosagecompensated in males. As a consequence, the transheterozygous males produce twice as mudh GAL4 activity as the femabes, and die during larval stages, whereas the femabs survive. Thus, all cutides shown are derived from females.
15. The full-bngth embryonic cDNA was reconstructed in a Bluescript KS+ backbone from three Eco Rl fragments. The full-bngth embryonic cDNA begins with a Hind lil site tat position 45 in the published sequence (3)] and ends with an Xba I site (constructed by inserting Xba l linkers in the Msl I site at position 2741). The cDNA was inserted as an Xho WXba I fragment into the GAL UAS vector [pUAST (11)]. This construction results in an oriented insertion in which the cDNA is proceeded at the 5' end by five optimized GAL4 binding sites, an hsp70 TATA box, the transcriptional start, and the cDNA is folbwed at the 3' end by the SV40 intron and polyadenylation site. A y as w stock was transfomed as described [G. M. Rubin and A. C. Spradling, Science 218, 348 (1982)]. A total of 13 independent pUAST ey strains were analyzed. The heat-inducible construct was made by inserting the embryonic d)NA into the heat-shock Casper vector [V. Pirrotta, in A Survey of Magyar Chants Vectors and 77;teir Uses, R. L. Rodriguez and D. T. Denhardt (Buttesworth, Boston and London,1988), p. 437].
16. G. M. Edelmann,Annu. Rev. CellBiol. 2, 81 (1986).
17. Wild-type and ectopic eyes were dissected and fixed for 30 minutes on ice in a cacodylate-buffered glutaraldehyde-oxmiumtetroxide fixation mixture. After postfixation in cacodylate-buffered osmiumtetroxide, the tissue was dehydrated through an ethanol series and embedded in Spurr medium. One-micrometer sections were cut and stained wrth staining solution (equal volumes of 2 percent Azur II and 2 percent disodiumtetraborate to 2 percent methylene blue). After drying they were mounted with Depex.
18. Staining of imaginal discs wrth ELAV antibodies was perfommed according to S. Robinow and K. White [J. Neurobiol. 22,443 (1991)]. A fluorescein-conjugateJ secondary rat antibody (Cappel) was used. Analysis was done on a Zeiss Axiophot microscope equipped for epifluorescence.
19. 1. A. Meinertzhagen and T. E. Hanson, in 77te Development of Drosophila melangoaster, M. Bate and A. Martinez-Arias, Eds. (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1993), chap. 24, p.
20. P. Callaerts, G. Halder, W. Gehnng, in preparation.
21. G. M. Rubin, personal communication.
22. L. M. Hall, P. J. Mason, P. Spierer, J. Mol. Biol. 169, 83 (1983); B. Bossy, L. M. C. Hall, P. Spierer, EMBO 3,2537 (1984); J. Gauze L. M. C. Hall, A. Spierer, P. Spierer, Genetés 112, 65 (1986); J. MirkovHch, P. Spierer, U. K. Laemmli, J. Mol. Biol. 190, 255 (1986). The haploid genome size of Drosopttib is estimated to be 165,000 kb. The gene density can be estimated from the number of transcription units detected on a chromosome walk of 315 kb and amounts to approximately one gene per 7 kb. After subtraction of 22 percent of satellite and other repetitive DNA, we obtain a value of approximately 17,000 genes per haploid genome. This is certainly an underestimate because not all transcripts from this region have been detected, and on the basis of expenence wHh genomic sequencing of yeast chromosomes, a considerably larger value is to be expected.
23. B. N . R. Cheyette et al. , Neuron 12,977 (1994); M. A. Serikaku and J. E. O'Tousa, Genetics 138, 1137 (1994).
24. B. Dickson and E. Haten, in 71&laqno;e Development of Drosophila melangoaster, M. Bate and A. MartinezArias, Eds. (Cold Spring Harbor Laboratory Press Cold Spring Harbor, NY, 1993), chap. 23, p. 1327, S. L. Zipursky and G. M . Rubin, Annu. Rev. NeLtrosci. 17, 373 (1994).
25. H. Spemann, Embryonic Devebpment and Induc-; tion (Yale University Press, New Haven, CT,1938). |
26. The full-length mouse Pax-6 cDNA (a gift of C. Walher and P. Grass) was doned as a Not WXho I Eminent in the GAL-UAS vector [pUAST (11)]. Flies were transfommed as described in (15). To ectopically induce the mouse Pax-6 gene, the UAS-Pax-6 transfommant lines were crossed to the E132 GAL4 expressing line as for the Drosopitib gene (14). The figure shows an ectopic eye on a second leg of a male.
27. J. Malicki et al. , Cell 63,961 (1990); J. J. Zhao, R. A. Lazzarini, L. Pick, Genes Dev. 7, 343 (1993); D. Bachilbr et al. , EMBO J. 13,1930 (1994).
28. ß-Galactosidase staining was perfommed as descnbed in M. Ashbumer, Drosophila, A Laboratory Manual (Cold Spnng Harbor Laboratory Press, Cold Spnng Harbor, NY,1989), protocol 77.
29. For cutide preparatior!s, adults were dissected in phosphate-buffered saline, mounted in Hoyer's or Faure's mounting medium, and immediately photographad to avoid diffusion and bleadhing of eye pigments.
30. For scanning electron microscopy, freshly hatched flies were narcotszed and immersed in 70 pernt atone. After critical point drying, they were mounted and coated with gold. The specimens were observed with a Hitachi S-800 field emission electron microscope at 6 kV.
31. We thank the following colleagues for fly stodes, antibodies, and plasmid vectors: S. Benzer, A Brand, C. Goodman, P. Gnuss, E. Haven, F. Jimenez, W. E. Kalisch, C. O'Kane, V. Pirrotta, C. Walther, and L Zipursky; C. O'Kane and his group for the support of one of us during his stay in the O'Kane laboratory (G.H.); E. Haten and collaborators for advice on histological and antibody staining techniques (to P.C.); and the Interdepartmental Electron Microscopy Servi for assistant in particular A Hetti and U. Sauder; E. Marquardt-Wenger for processing the manuscript; and M. Affolter, W. Keegan, and F. Lossli for critical discussions. Supported by the Colon Foundation of S Leuven, Belgium (P.C.), the Kantons of Basel, and the Swiss National Science Foundation.
1 February 1995; accepted 24 February 1995
Return To Molecular Biology Main Page
![]()
© Copyright
2000 Department of Biology, Davidson College, Davidson, NC 28036
Send comments, questions, and suggestions to: macampbell@davidson.edu