Figure 1
Structure of the Hrdc domain of the SGS protein
Figure taken from the PDB database: http://www.rcsb.org/pdb/index.html
PDB ID: 1D8B
Primary citation Liu et al.,
1999.
MORE INFO
SUPPRESSION OF ILLEGITAMATE
RECOMBINATION
In sgs1 mutants an increase in illegitimate recombination
via the RAD52 and Hdf1 homologous recombination pathway is observed ( Yamagata,
1998). This suggests that sgs1 suppresses illegitimate recombination
by regulating the Rad52 and Hdf1 homologous recombination pathways (Yamagata,
1998).
RNA POLYMERASE II TRANSCRIPTION
Defects in the synthesis of RNAII polymerase
transcripts were observed in sgs1, srs1 double mutants (Lee,
1999). This suggests that sgs1 and srs2 interact with RNA polymerase
II. Lee et al. (1999) propose that srs1 and sgs1 help DNA unwind
during RNA polymerase II transcription. When srs1 and sgs1 are not
present, the DNA does not unwind as rapidly. Failure to unwind causes
the RNA polymerase to pause which results in double stranded breaks (DSB)
that are fixed by homologous recombination. Fixing DSB with homologous
recombination would account for the deletions seen in the rDNA.
INTERACTION WITH TOPOISOMERASES
The N terminus of the sgs1 protein binds to
topoisomerase III (topIII) ( Bennett,
2001). Topoisomerases relieve the super coiling found during DNA
replication, due to the unwinding of the DNA helicases, by snipping and
later rejoining super coiled DNA (Griffiths,
1999).
Cells with mutant sgs1 copies also show an increase
in chromosome missegreation (Watt,
1995). SGS1 interacts with topII during chromosome segregation (Watt,
1995).
PREMATURE DEATH-CELL CYCLE CHECK
POINTS
SGS1 also plays a role in cell cycle checkpoints. Frei
et al. (2000) suggest that sgs1 interacts upstream of Rad53 in the S cell
cycle check point. They suggest that sgs1 normally halts the progression
past the S stage when there is a stalled replication fork. Mutant
sgs1 yeast sometimes fail to activate the S checkpoint when there is a
stalled replication fork.
McVey et al. (2001) suggest
that the early cell death of sgs1 mutants can be attributed to two causes.
One is the arrest of the cell cycle at the G2/M checkpoint and the other
is an arrest in the G1 stage due to causes seen in normal age related senesence.
McVey et al. (2001) propose that when sgs1 is not present, the cell causes
DSBs and then uses homologous recombination to fix stalled replication
forks. When the DSBs or the homologous recombination complex can
not be resolved the cell cycle is arrested in the G2/M check point.
When this happens the cell dies as a small bud is beginning to come off
the mother cell. Occasionally the cell overlooks the DNA damage and
continues into mitosis. When this happens the cells die in the next
few generations due to the irreparable damage caused by the DSBs.
When cells arrest in the G1 stage it is usually
accompanied by fragmentation of the nucleolus ( Guarente,
1997) and an accumulation of extra chromosomal regions (ERC) (McVey,
2001). In sgsI mutants this seems to occur about 60% sooner
than in wild type cells (Guarente, 1997).
This is thought to be the result of the hyperrecombination and increased
homologous recombination seen in sgs1 mutants ( McVey,
2001).
BLOOMS SYNDROME
Blooms syndrome is a human disease caused
by mutations in the BLM gene ( Watt,
1996). Mutations in the BLM gene result in growth retardation, increased
incidence of cancer, and genomic instability (Watt,
1996). Unlike the other homologous genes to BLM, BLM and SGS1 share
a highly charged N terminus (Watt, 1996). SGS1
mutants are being used to model blooms disease in yeast.
For a picture of homologous sections between the bloom's syndrome gene
and sgs1 go to Figure 1 of the following article by Watt et al. (1996):
http://journals.bmn.com/journals/list/render?uid=JCUB.bb6308&node=TOC%40%40JCUB%4011%4018%4011_18
WERNERS SYNDROME
Werners syndrome is a human disease caused
by a mutation in the WRN gene (Guarente, 1997).
The main characteristic of Werners syndrome is premature aging (Guarente,
1997). SGS1 is homologous to the WRN gene and thus is also being
used as way to model Werners disease (Guarente,
1997).
UNANNOTATED GENE
BASIC INFO
SGD: http://genome-www4.stanford.edu/cgi-bin/SGD/locus.pl?locus=YJU3
Id: gene: YJU3; ORF: YK1094W
Swiss Port: http://www.expasy.ch/cgi-bin/niceprot.pl?P28321
Id: P287321
YPD: http://www.proteome.com/databases/YPD/reports/YJU3.html
Genbank Id: CAA81932
Name: YJU3
Chromosome: 11
Null allele: viable
DNA sequence:
1 ATGGCTCCGT ATCCATACAA
AGTGCAGACG ACAGTACCTG AACTTCAATA
51 CGAAAACTTT GATGGTGCTA AGTTCGGGTA CATGTTCTGG CCTGTTCAAA
101 ATGGCACCAA TGAGGTCAGA GGTAGAGTTT TACTGATTCA TGGGTTTGGC
151 GAGTACACAA AGATTCAATT CCGGCTTATG GACCACTTAT CACTCAATGG
201 TTACGAGTCA TTTACGTTTG ATCAAAGGGG TGCTGGTGTT ACATCGCCGG
251 GCAGATCGAA AGGTGTAACT GATGAGTACC ATGTGTTTAA CGATCTTGAG
301 CATTTTGTGG AGAAGAACTT GAGTGAATGT AAGGCCAAAG GCATACCCTT
351 GTTCATGTGG GGGCATTCAA TGGGCGGTGG TATCTGCCTA AACTATGCCT
401 GCCAAGGTAA GCACAAAAAC GAAATAAGCG GATATATCGG GTCAGGCCCA
451 TTAATAATTT TACATCCGCA TACAATGTAT AACAAGCCGA CCCAAATTAT
501 TGCTCCATTA TTGGCGAAAT TTTTACCAAG GGTAAGGATC GACACTGGTT
551 TAGATCTTAA AGGAATCACA TCTGATAAAG CCTATCGTGC TTTCCTCGGA
601 AGCGATCCTA TGTCTGTTCC ACTATATGGG TCGTTTAGGC AAATACACGA
651 CTTTATGCAA CGTGGTGCCA AGCTCTACAA GAATGAAAAC AATTATATTC
701 AGAAGAACTT CGCTAAAGAC AAACCCGTTA TTATTATGCA TGGACAAGAC
751 GACACAATCA ACGATCCTAA GGGCTCTGAA AAGTTCATTC AGGACTGTCC
801 TTCTGCTGAC AAAGAATTAA AGCTGTATCC GGGCGCAAGA CATTCGATTT
851 TCTCACTAGA GACAGATAAA GTCTTCAACA CGGTGTTCAA TGATATGAAG
901 CAATGGTTGG ACAAACACAC CACGACCGAA GCTAAACCAT AA
Protein sequence:
1 MAPYPYKVQT TVPELQYENF
DGAKFGYMFW PVQNGTNEVR GRVLLIHGFG
51 EYTKIQFRLM DHLSLNGYES FTFDQRGAGV TSPGRSKGVT DEYHVFNDLE
101 HFVEKNLSEC KAKGIPLFMW GHSMGGGICL NYACQGKHKN EISGYIGSGP
151 LIILHPHTMY NKPTQIIAPL LAKFLPRVRI DTGLDLKGIT SDKAYRAFLG
201 SDPMSVPLYG SFRQIHDFMQ RGAKLYKNEN NYIQKNFAKD KPVIIMHGQD
251 DTINDPKGSE KFIQDCPSAD KELKLYPGAR HSIFSLETDK VFNTVFNDMK
301 QWLDKHTTTE AKP
Conserved Domain Search
http://www.ncbi.nlm.nih.gov/Structure/cdd/cddsrv.cgi?uid=pfam00561&version=v1.54
pfam 00561
abhydrolase, alph/beta hydrolase fold
catalytic domain found in many enzymes
ProDom
http://protein.toulouse.inra.fr/cgi-bin/ReqProdomII.pl?acc_seq0=P28321
yeast-
complete proteome peroxiase lysophospholipase chloroperoxidase synthase
polymerase
oxidoreductase non-heme
homologous proteins-
function: biotransformation enzyme that catalyzes the hydrolysis of
epoxides (alkene oxides,
oxiranes) and arene oxides to less reactive and
more water soluble dihydrodiols by the trans
addition of water
catalytic activity: epoxide + H20 = glycol
NCBI BLASTP
http://www.ncbi.nlm.nih.gov/BLAST/
abhydrolase
40-50% positives with lisophospholipases of other organisms
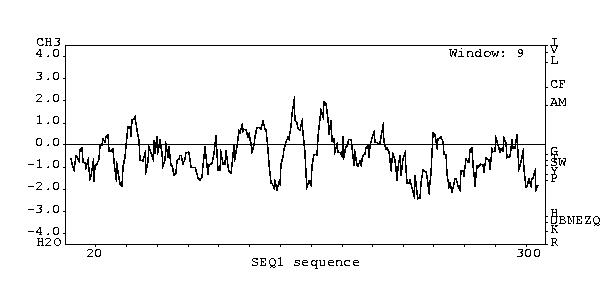
Kyte Doolittle Plot
http://fasta.bioch.virginia.edu/fasta/grease.htm
J. Kyte and R. F. Doolittle (1982) J. Mol. Biol. 157:105-132
The Kyte Doolittle hydropathy plot tells you whether a protein may me
a transmembrane protein.
If a peak is higher than two then the protein may be a transmembrane
protein.
Figure 2 Kyte Doolittle Hydropathy Plot for the sgs1 protein
 d
d
There doesn't seem to be very good evidence to suggest that the YJU3
gene encodes for a transmembrane protein.
BLASTP against other mamalian homologs
http://genome-www.stanford.edu/cgi-bin/SGD/Sacch3D/getblast?name=YJU3&db=mammal
Altschul, Stephen F., Warren Gish, Webb Miller, Eugene W. Myers, and
David J. Lipman (1990).
Basic local alignment search tool. J. Mol. Biol.
215: 403-10.
Altschul et al. (1997), Gapped BLAST and PSI-BLAST: a new generation
of protein database
search programs. Nucl. Acids Res. 25: 3389-3402.
46% positive with human lysophospholipase homolog
44% positive with Mouse cyclophilin C-associated protein
44% positive with Mouse mama gene product
PULLING IT ALL TOGETHER
The data suggest that YJU3 may be a abhydrolase
protein and a lysophospholipase protein. Experiments should be created
and performed to test this possibility.
References
Bennett, Richard J.
and James C. Wang. September 25, 2001. Association of yeast DNA
topoisomerase III and Sgs1 DNA helicase: Studies
of fusion proteins. PNAS (USA) 98(20):
11108-11113. http://www.jbc.org/cgi/content/abstract/275/35/26898?ijkey=12NNavZs9ppNo
Frei, Christian and Susan M.
Gasser. January 2000. The yeast Sgs1p helicase acts upstream of
Rad53p in the DNA replication checkpoint and colocalizes
with Rad53p in S-phase-specific
foci. Genes and Dev. 14(1): 81-96.http://www.genesdev.org/cgi/content/full/14/1/81
Griffiths, Anthony,
W.M. Gelbart, J.H. Miller, R.C. Lewontin. 1999. Modern Genetic
Analysis.W.H. Freeman and Company, New York,
pp. 88-90.
Guarente, Leonard.
October 1997. Link between aging and the nucleolus. Genes and Dev. 11(19):
2449-2455. http://www.genesdev.org/cgi/content/full/11/19/2449
Lee, S. K. , Johnson, R. E. ,
Yu, S. L. , Prakash, L. & Prakash, S. 1999. Requirement of Yeast
SGS1 and SRS2 genes for replication and transcription.
Science
286: 2339-2342.
http://www.sciencemag.org/cgi/content/full/286/5448/2339?ijkey=bWVP.CI6.mh6A
Liu, Z., Macias, M. J., Bottomley,
M. J., Stier, G., Linge, J. P., Nilges, M., Bork, P., Sattler, M. 1999.
The Three-Dimensional Structure of the Hrdc Domain and Implications for
the Werner
and Bloom Syndrome Proteins. Structure (London)
7: 1557.
McVey, M. , Kaeberlein, M.
, Tissenbaum, H. A. & Guarente, L. 2001. The short life span of
Saccharomyces servisiae sgs1 and srs2 mutants is
a composite of normal aging processes and
mitotic arrest due to defective recombination. Genetics
157: 1531-1542.
http://www.genetics.org/cgi/content/full/157/4/1531
SGD database. 2001.Stanford.
http://genome-www4.stanford.edu/cgi-bin/SGD/locus.pl?locus=YJU3
Swiss-Port. 2001. http://www.expasy.ch/cgi-bin/niceport.pl?P35187
Watt, Paul M. and Ian D. Hickson.
1996. Failure to unwind causes cancer.
Current Biology.
6:265-267. http://journals.bmn.com/journals/list/browse?uid=JCUB.bb6308&rendertype=text
Watt PM, Louis EJ, Borts
RH, Hickson ID. April 1995. Sgs1: a eukaryotic homolog of E. coli
RecQ that interacts with topoisomerase II in vivo
and is required for faithful chromosome
segregation. Cell. 81(2): 253-60.
Yamagata K, Kato J,
Shimamoto A, Goto M, Furuichi Y, Ikeda H. July 1998. Bloom's and
Werner's syndrome genes suppress hyperrecombination
in yeast sgs1 mutant: implication for
genomic instability in human diseases. PNAS (U S
A) 95(15):8733-8.
http://www.pnas.org/cgi/content/full/95/15/8733
YPD database. 2001. Proteome,
Inc.
http://www.proteome.com/databases/YPD/reports/YJU3.html
Contact info: lirobinson@davidson.edu