This web page was produced as an assignment for an undergraduate course at Davidson College.
Gln3 Protein
Gln3 protein is a transcription factor found in the nucleus and cytoplasm which plays a role in nitrogen catabolite repression (NCR). It binds the nitrogen upstream activation sequence of Gln1, the gene encoding glutamine synthetase (Swiss-Prot, 2002, http://www.expasy.org/sprot/).
The amino acid sequence for Gln3 obtained from NCBI (NCBI, 2002, http://www.ncbi.nlm.nih.gov/ )
1 mqddpenskl ydllnshldv hgrsneeprq tgdsrsqssg ntgeneedia fasglnggtf
61 dsmlealpdd lyftdfvspf taaattsvtt ktvkdttpat nhmdddiamf dslattqpid
121 iaasnqqnge iaqlwdfnvd qfnmtpsnss gsatisapns ftsdipqynh gslgnsvsks
181 slfpynssts nsninqpsin nnsntnaqsh hsfniyklqn nnssssamni tnnnnsnnsn
241 iqhpflkksd siglsssntt nsvrknslik pmsstslanf kraasvsssi snmepsgqnk
301 kpliqcfnck tfktplwrrs pegntlcnac glfqklhgtm rplslksdvi kkriskkrak
361 qtdpniaqnt psapatasts vtttnakpir srkkslqqns lsrvipeeii rdnigntnni
421 lnvnrggynf nsvpspvlmn sqsynssnan fngasnanln snnlmrhnsn tvtpnfrrss
481 rrsstssnts sssksssrsv vpilpkpspn sansqqfnmn mnlmnttnnv sagnsvassp
541 riissanfns nsplqqnlls nsfqrqgmni prrkmsrnas ysssfmaasl qqlheqqqvd
601 vnsntntnsn rqnwnssnsv stnsrssnfv sqkpnfdifn tpvdspsvsr pssrkshtsl
661 lsqqlqnses nsfisnhkfn nrlssdstsp ikyeadvsag gkisednstk gsskessaia
721 deldwlkfgi
DIP (Database of Interacting Proteins, 2002)
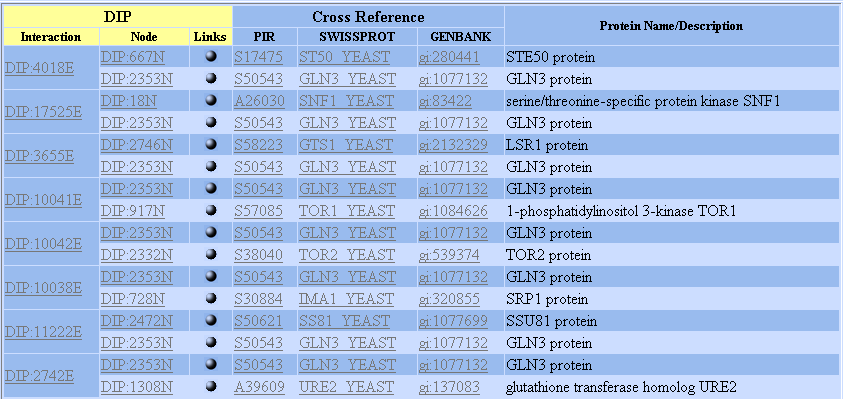
Below you can see that Gln3 is connected to many different proteins. I will list the functions of these proteins below the chart.
STE50 protein is involved in growth arrest during conjugation and may interact with the G protein alpha subunit. (Swiss-Prot, 2002, http://www.expasy.org/sprot/) Interaction with Gln3 supported by a yeast two hybrid test; Nature 2000 Feb 10;403(6770):623-7.
SNF1 protein is essential for release from glucose repression.(Swiss-Prot, 2002, http://www.expasy.org/sprot/) Interaction with Gln3 supported by yeast two hybrid experiment [Mol Cell Biol 2002 Feb;22(4):1246-52t], affinity column and two link validation tests (small scale, non-genome wide experiments).
LSR1 appears to regulate timing of budding to obtain appropriate cell size independent of DNA replication.It is a transcription factor involved also in heat resistance and flocculation (Swiss-Prot, 2002, http://www.expasy.org/sprot/). Interaction with Gln3 supported by yeast two hybrid experiment [Nature 2000 Feb 10;403(6770):623-7].
TOR1/TOR2 are phosphatidylinositol 3-kinase homologs required for G1 progression. They are targets for antibiotic Rapamycin..(Swiss-Prot, 2002, http://www.expasy.org/sprot/). Interaction with Gln3 supported by yeast two hybrid [J Biol Chem 2000 Nov 17;275(46):35727-33], affinity column, and three link validation tests.
SRP1 protein binds specifically and directly to substrates with either a simple or bipartite NLS motif. It promotes docking of import substrates to nuclear envelope.(Swiss-Prot, 2002, http://www.expasy.org/sprot/). Interaction with Gln3 supported by immunoprecipitation [J Biol Chem 2001 Jul 6;276(27):25359-65] and one link validation test.
SSU81 protein interacts with RNA Polymerase II. It might induce PB2P autophosphorylation and is a potential integral membrane protein (Swiss-Prot, 2002, http://www.expasy.org/sprot/). Interaction with Gln3 supported by yeast two hybrid [Science 2002 Jan 11;295(5553):321-4].
URE2 protein plays an important role in response to nitrogen source. It helps repress Gln1 and Gdh2 and interacts with glutamine synthase. URE2 may act as a catalyst to inactivate Gln3 in response to high levels of intracellular glutamine. (Swiss-Prot, 2002, http://www.expasy.org/sprot/). Interaction with Gln3 supported by immunoprecipitation [J Bacteriol 1996 Aug;178(15):4734-6], affinity column [Nature 1999 Dec 9;402(6762):689-92], yeast two hybrid test [J Biol Chem 2001 Aug 24;276(34):32136-44], and two link validation tests.
All of these proteins appear to have roles important in metabolism. Aside from SSU81, which is a potential integral membrane protein, they seem to be located in the nucleus. Below is a protein network. The red dot in the center is Gln3. As you can see, Gln3 is highly connected to many other proteins.
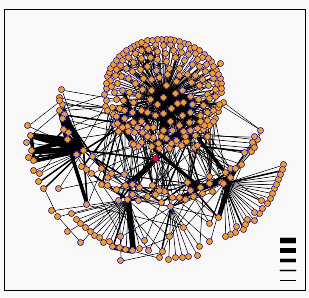
Image from DIP database, 2002.

If you clicked on TN7-20F2 (the one above that says out-of frame), you come up with some interesting information.

This page shows that when Tn3, a transposon at first used to locate ORF's, found a potential NORF at codon 4 of Gln3. NORF stands for nonannotated ORF's and are often passed over because they are <100amino acids and are hidden within larger ORF's. Since Gln3 is a rather large protein, 730 amino acids, it is not shocking that another ORF could be hidden with its DNA coding.
YRC (Yeast Resource Center)
Yeast two hybrid method uses bait proteins to catch prey proteins. Prey protein are proteins that will interact and bind to bait proteins. The YRC lists and links many proteins which interact. However, my protein was listed under new two-hybrid interactions generated since last survey published in Nature 403: 623-627.
Bait protein and prey protein
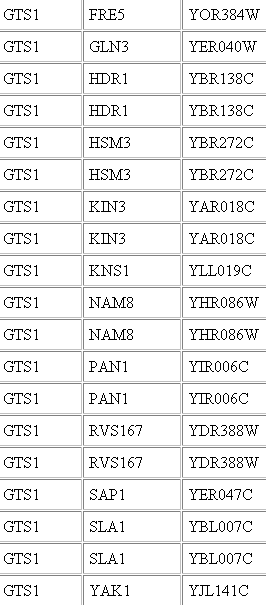
These are just a few of the proteins which were baited by GTS1. Gln3 is second from the top.

Thus it appears GTS1 bind to each other Gln3. It seems likely since GTS1 plays a role in the cell cycle, as did most of the other proteins which were listed attached to Gln3 in DIP.
Protein circuit diagram produced by Benno Schwikowski at the Institute for Systems Biology. Gln3 is in the center.
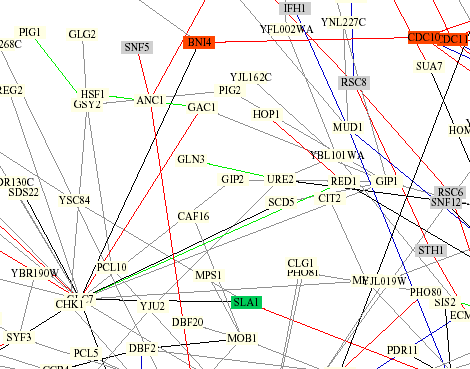
It appears that Gln3 is only connected to URE2 by a green line. This indicates their cellular roles are identical but they are located in different places. The light yellow box around Gln3 means unknown role, which seems strange since the function of Gln3 is pretty much figured out.
No interactions shown for Gln3 in Function Junction.
MIPS had a lot of information on Gln3. It describes it as a transcription factor for positive nitrogen regulation. It also says "the four GATA factors Gtz3p, Dal80p, Gln3p, and Gat1 regulate each other and the expression of nitrogen catabolic genes" [J Bacteriol. 1997 Jun;179(11):3416-29.]


Conclusions: Gln3 interacts with a fair number of proteins in its regulation of nitrogen metabolism. The number is not overwhelming, yet we do not know the whole extent of its interactions. This is just the beginning to understanding the role these genes play in nitrogen metabolism. The questions that remain include when, where, and if Gln3 and the other proteins are together at the same points in time. Do they all bind at once or at separate intervals? This is the future of proteomic research.
YEN1 Protein
MIPS describes YEN1 as having a weak similarity to DNA repair protein Rad2p and Dsh1p.
DIP
No information found for YEN1.
No information found for YEN1.
No information found for YEN1.
No interactions shown for YEN1 in Function Junction.
Briefly calls YEN1 a hypothetical 87.4 kDa protein in Gln3-SAH1 intergenic region.
Future Experiments
Gln3 expression in different types of nitrogen sources has been pretty well catalogued. However, how the Gln3 protein is activated in different levels of nitrogen has not been fully discussed. Using the ICAT method, we could compare GLN3 in low levels of nitrogen vs. high levels of nitrogen. Using a 2D gel and then MS to isolate Gln3 from the different cells, we could label it with antibodies specific for phosphoproteins to see if Gln3 is phosphorylated in response to environmental nitrogen levels. If Gln3 shows different levels of phosphorylation, we could perform a Y2H test using the phosphorylated and unphosphorylated version pf Gln3 as the bait protein. We could see what effect phosphorylation has with respect to the specificity of Gln3 to binding proteins. The MIPS database said that Gtz3p, Dal80p, Gln3p, and Gat1 regulate each other. Using RNA interference (RNAi) for each protein individually, we could study the pathway of interaction with these four genes. So, if you knock out one protein, will the other three still interact, will the pathway be broken, what other genes might be induced (microarray or gas chromatography and MS to study entire metabolome with mutant of one of these genes and wild-type), can the cell still metabolize nitrogen? All these things can help understand the pathway to nitrogen metabolic regulators.
Before I had predicted that YEN1 had something to structural elements in the cell cycle or (according to the microarray of expression in response to histone depletion for YEN1) a DNA housekeeping gene. This concurs with MIPS saying YEN1 has weak similarity to DNA repair proteins. For YEN1 I would begin by performing an immunofluorescence test to see exactly where the protein is located within the cell. If it is in the nucleus, it being a DNA repair protein would be supported. If in the cytosol, the structural hypothesis would seem more likely.
What might be the most fruitful preliminary step after determining locale would be to use YEN1 as the bait protein in a Y2H experiment. Scanning many proteins for YEN1 interactions would give helpful insight into its cellular role and what pathways it is involved in. If YEN1 is a DNA repair protein, an experiment could be performed which quantified with ICAT and MS if there were different levels of YEN1 in cells that were exposed to increasingly higher levels of radiation. If we see the levels of YEN1 protein rise in response to DNA damaging radiation, we could predict that YEN1 had something to do with DNA repair. However, we would not know if the radiation had other effects on cell function, but its a start.
If we discover what proteins YEN1 actually interacted with, we would have a much stronger basis with which to design experiments.
Questions or comments? Email me: laquillian@davidson.edu
© Copyright 2003 Department of Biology, Davidson College, Davidson, NC 28035