
This web page was produced as an assignment for an undergraduate course at Davidson College
My Favorite Yeast Proteins: CYT1 and YOR066W
| The SGD database identifies roles regarding the biological processes, molecular functions and cellular component of the Cyt1 encoded protein. While data support Cyt1's role as a mitochondrial electron transport cytochrome, there is much about this protein that Gene Ontology (GO) annotations do not tell us. Because it is non-annotated, GO tells us even less about the gene product of YOR066W. The molecular function, biological process and cellular component roles of YORR066W are, as of now, unknown, but the results of a yeast two hybrid study (Y2H) indicate that YOR066W is potentially a substrate for Cdc28 or a cyclin protein (SGD, 2004). Definitive predictions ascertaining the role of YOR066W in yeast cells have not been found in previous web pages which investigated the YOR066W gene and its expression pattern. Trend comparisons of induction and repression in Cyt1 and YOR066W are sometimes similar, as in the case of histone depletion, and sometimes dissimilar, as in the context of diuxic shift. Clustered microarray data group similarly transcribed genes but do not necessarily determine groupings of similarly functioning proteins. Here, we will use the tools that may further elucidate the functions of Cyt1 and YOR066W. |
CYT1 and Y2H
The yeast two-hybrid (Y2H) system is a method that uses a particular protein to discover proteins with which that protein interacts (Campbell, 2003).
Figure 1. This screen shot shows no protein interactions between Cyt1 and another protein. Since proteins sometimes bind one way, but not the other (i.e. RPC19 binds to bait RPC40 but RPC40 does not bind to prey RPC19), I checked to see if Cyt1 interactions were detected when Cyt1 was listed as prey, instead of bait. There were no protein interactions when CYT1 was used as the activation-domain-bound prey. |
This image makes interactions between the bait Cyt1 and the ORFs of other yeast genes in the Y2H method appear absent, assuming a genomic library was transformed into plasmids where CYT1 was being used as bait, CYT1 does not appear to interact with any other yeast proteins. Since the interactions detected using this method must occur inside the nucleus of the yeast cell, and CYT1 is located in the mitochondria, it is possible that CYT1 does not work well inside the nucleus. A conformational change in structure could cause a misshapen protein which, inside the nucleus, does not bind to the proteins with which we might expect CYT1 to interact. CYT1 as a "prey" protein might also be misshapen and thus behave differently inside the nucleus than in the mitochondria. Another possibility for the lack of interaction is that the Y2H experiment has yet to be run with Cyt1. |
CYT1 and DIP
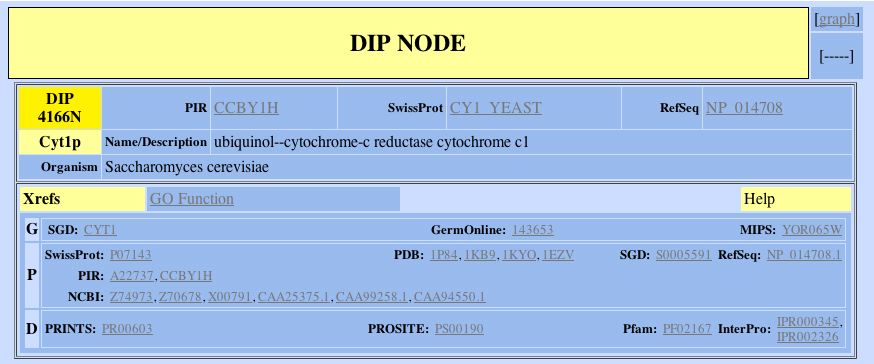
Figure 2. This shows the one hit for CYT1 from the DIP database. Note the various terms used to refer to the same protein in different databases.
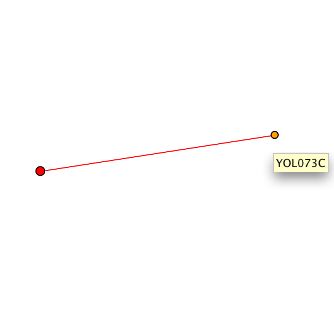 |
Figure 3. The red node on the left is the CYT1 protein. One interaction, with protein YOL073C appears. Note that this is one more protein than was found to interact using the Y2H method. |
 |
Figure 3. The protein that appears to interact with the CYT1 protein in the graph above shows no indication of known function, process or cellular component (as indicated by "ND"). At the time of annotation, there were no publications for this gene (SGD, 2002). Note that this hypothetical ORF occurs upstream of, on the same chromosome as CYT1. |
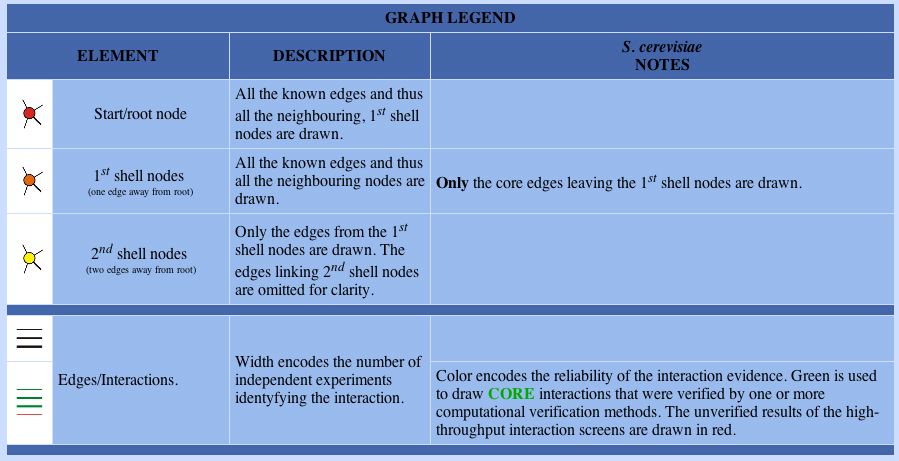 |
Figure 4. The explanation of the root and 1st shell nodes seen in the map, above, are color coded and described, here.
| These figures disagree with the results from the Y2H experiment. While I would have expected the graphed web of proteins, which interact with the CYT1 protein, to involve mitochondrial proteins and cytochrome gene product proteins, the DIP database shows interaction with only the non-annotated ORF, YOL073C, located on the same chromosome as CYT1. |
CYT1 and PDB
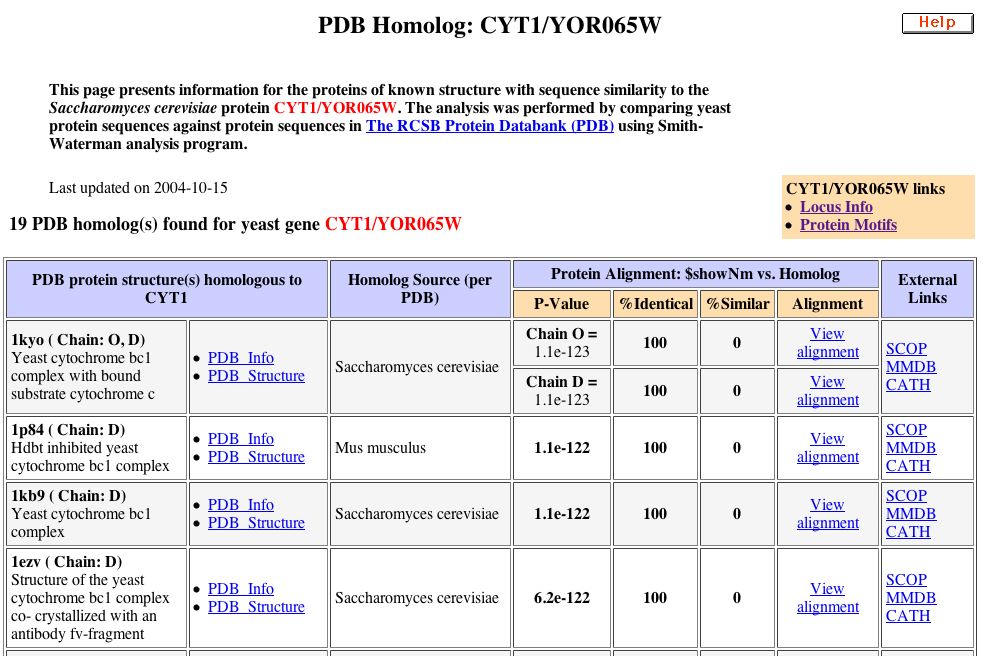
Figure 5. Five of the nineteen homologs found for CYT1 were 100% identical (SGD, 2004).
Figure 6. This is what the EZ1 homolog of the CYT1 protein looks like (PDB, 2004).
| This protein is conserved in several species; homologs exist in mouse, chicken and cow. It is not surprising that CYT1 homolog proteins are evident in other species since mitochondrial electron transport is a biological process that occurs in these species, as well. |
CYT1 and KEGG
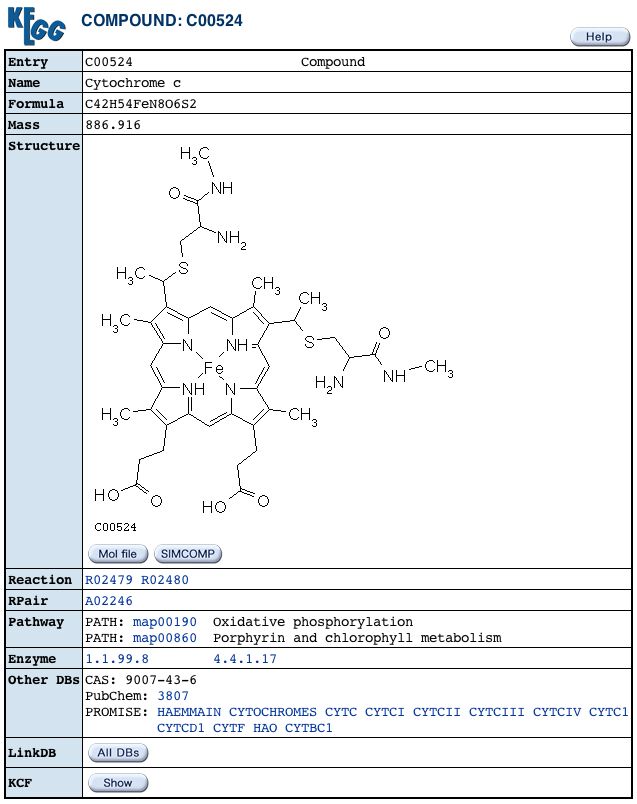 |
Figure 7. Here we see the chemical properties of formula, mass and structure for cytochrome c, the compound to which Cyt1 transports electrons (KEGG, 2004). |
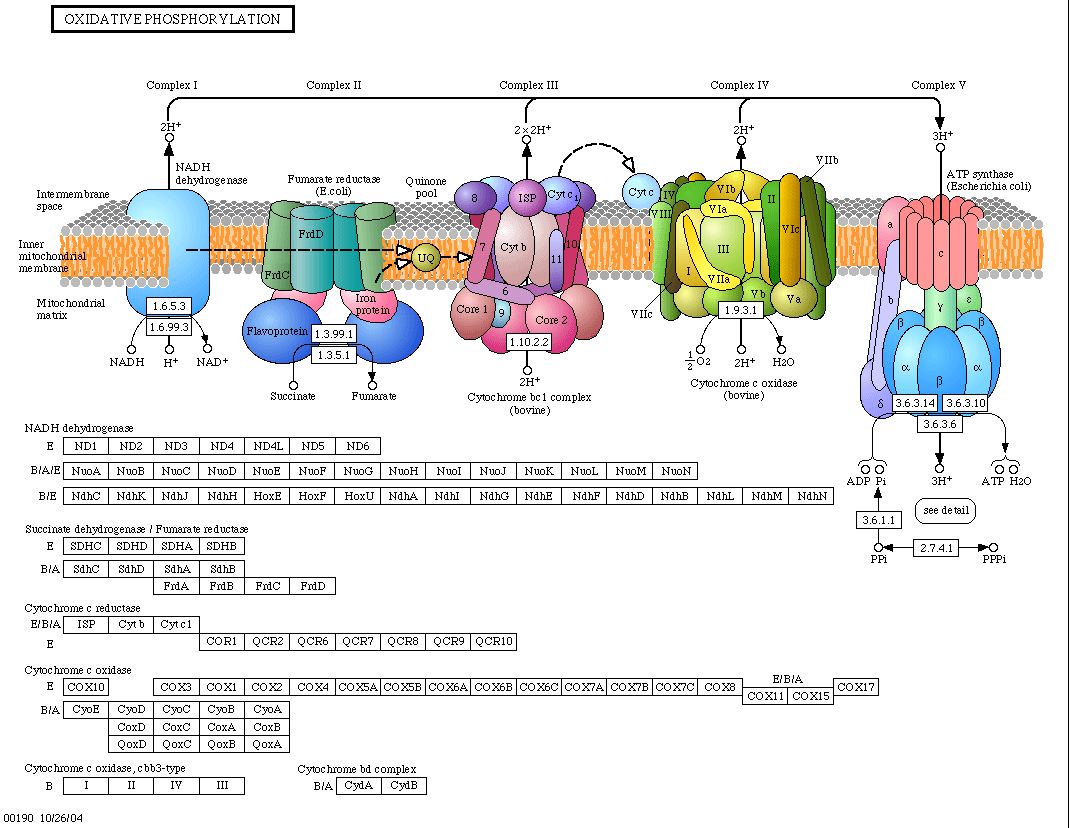 |
| Figure 8. Here we see cartoon interactions of the proteins involved in oxidative phosphorylation. Note that CYT1 (cytochrome c is labeled Cyt c, in this diagram) interacts with the proteins of the inner mitochondrial membrane and proteins in cytochrome c oxidase. The boxes below this cartoon categorize the proteins involved in each aspect of oxidative phosphorylation. Note that Cyt c1 is in the Cytochrome c reductase category (KEGG, 2004). |
| We are confronted again with the conservation that occurs in biologically imperative processes. In this illustration of oxidative phosphorylation, E. coli and bovine protein complexes alternate to give us an idea of what proteins and mechanisms are required for oxidative phosphorylation and the similarity between bacteria and cow oxidative phosphorylation. This diagram shows us the circuitry that most likely applies to CYT1, but it also acknowledges that there may also be proteins involved in this process that are yet to be discovered, represented by "IV" and "VIII" in complex IV, above, which interact with CYT1. transports to cyt. c. |
CYT1 and MIPS
 |
 |
Figure 9. The evidence that other yeast proteins are involved in the same biological process of electron transport, molecular function of oxdative phosphorylation and the same cellular component of inner mitochondrial membrane is well supported. There are over 1000 proteins involved in the cellular component of the mitochondria and over 100 of these are localized to the inner mitochondrial membrane (see right). There are also many other proteins involved in the same biological processes as Cyt1 (see left). |
| CYT1 is one of 1053 mitochondrial proteins, and one of 143 inner membrane mitochondrial proteins. there are still uncertainties regarding how it is sorted and where the intermediates of the protein exist as well as what interacts with the favorite protein and how. |
CYT1 and SGD
 |
| Figure 10. CYT1 is the only gene associated with the component aspect of the mitochondrial electron transport chain. CYT1 is one of several genes associated with the process aspect of mitochondrial electron transport from ubiquinol to cytochrome c (SGD, 2004). |
Conclusions about CYT1
In the 42 gene products associated with yeast cytochromes and the 55 gene products associated with yeast mitochondria (SGD, 2004), it is likely that many interactions with CYT1 protein occur. The data found from the DIP database do not describe the entire story of CYT1 protein interaction. There are at least two possible explanations for the fact that no data is exhibited in the interaction illustration of Cyt1 in the DIP database. Databases are being further developed constantly. DIP may not yet have tested the interactions between Cyt1 and all other yeast proteins. Another possibility is that Cyt1 has very limited interaction with other proteins, since Cyt1 serves as an electron transport molecule in the cytochrome c reductase complex. This could potentially mean that Cyt1 only interacts with one molecule which it transports from ubiquinol to cytochrome c. No results were found in the PDFs for membranes, degradation or aging proteins. I would have expected Cyt1 to appear in the membrane protein PDF, although perhaps inner mitochondrial membrane is structurally more similar to a membrane bound protein, than a membrane protein, in this case. Y2H protein interaction has probably not yet been investigated as there appear to be no interactions in this image and we can be fairly certain that Cyt1 interacts with at least one other protein in order to properly carry out its function. It is important to remember that Cyt1 is a mitochondrial protein and so it may not function, or interact, fully in the nucleus, which is where the Y2H method takes place. It is fairly clear, based on data from the SGD site, that this protein is involved in oxidative phosphorylation and located in the mitochondria of yeast. Investigations in proteomics show that there is much more to be figured out about the function of even relatively well understood proteins. |
YOR066W and Y2H
Bait |
Prey |
Prey ORF |
YOR066W |
YLR297W |
YLR297W |
Figure 11. There is a Y2H interaction between YOR006W and YLR297W proteins. (YRC, 2004)
Because the YOR066W protein hybridizes with the YLR297W protein we now have a new ORF to investigate. |
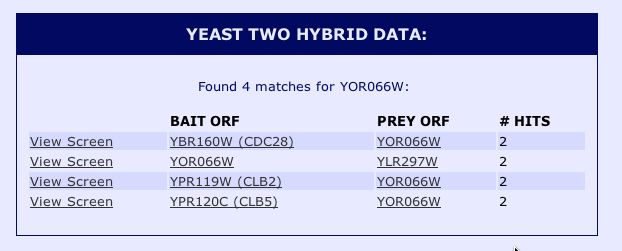
| Figure 12. While YOR066W only binds once when bound (as bait) to the DNA binding domain, this protein binds to several genes as the prey ORF (SGD, 2004). |
Interestingly, YOR066W does bind to CDC28 as a prey protein. There are two other proteins which are now of interest. CLB2 and CLB5 also bind to YOR066W in the Y2H experiment. There are interactions between YOR066W and known protein products. At this point we have three strong leads as to a potential function of YOR66W in its binding to Cdc28, Clb2 and Clb5. |
YOR066W and DIP

Figure 13. No protein interatcions were found from this YOR066W search (DIP, 2002).
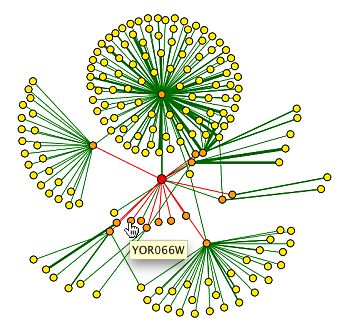 |
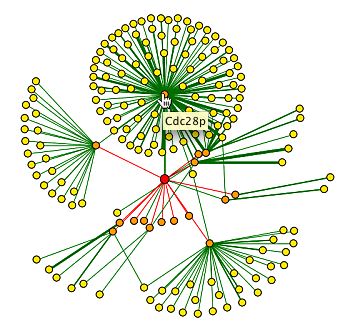 |
Figure 14. cyclin B5 is in the center. YOR066W is projected here as a 1st shell node. Cdc28 is also projected as a first shell node to Clb5 (DIP, 2002).
Given that there is no information regarding the direct interaction of YOR066W and other proteins, from the DIP database, it is useful to see interactions of known proteins with a non-annotated gene protein products. It is important to note that the design of this graph does not necessarily mean that there is no interaction between YOR066W and Cdc28. The red edge from Clb5 to YOR066W signifies that the interaction is from an unverrified, high throughput method. The green line between Clb5 and Cdc28 is better established and verified with computational methods. The data do not eliminate the possibility of more interaction of the YOR066W protein. |
YOR066W and PDB

Figure 15. There is no known structure for the YOR066W protein (PDB, 2004).
YOR066W and MIPS
 |
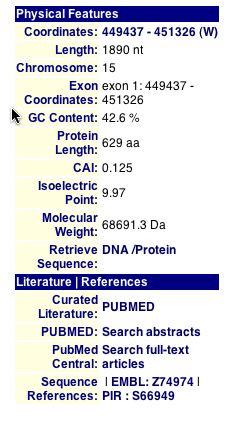 |
Figure16. Physical characteristics were found for YOR066W, using the MIPS database, but not much relevant information regarding its cellular or biological role (MIPS,2002).

Figure 17. If use the known proteins which were obtained in the earlier in the Y2H experiment, we see physical interactions between Clb5, Clb2 and Cdc28 (MIIPS, 2002).
CLB3, CLB5, CDC28 and SGD
| Clb2 |  |
 |
| Clb5 | |
 |
| Cdc28 | |
 |
| Figure 18. These three proteins all include a kinase activity, are involved in cellular cycle processes, and located in the cytoplasm and/or the nucleus. |
Conclusions about YOR066W
| While it is unlikely that the protein interactions of YOR066W is exclusive to those found to be Y2H bait, it is possible that YOR066W interacts with one or more of these proteins. From the protein interactions and the functional components of the known proteins with which YOR066W interacts, we can predict that YOR066W may play a role in yeast cell cycle. This is far from a complete analysis of the functional role of YOR066W, in fact it only questionably scratches the surface of functional role of this unknown protein. However, based on Y2H interactions and the role that bound proteins play in the yeast cell, we can resolve some of the potential functions of YOR066W and with each new protein, some of the larger issues of science become elucidated, as well. As we begin to better understand the interactions of genes and their protein products in yeast, we have a better chance of applying proteomics to such areas as drug development and disease diagnosis. |
Future Findings
Experiment 1: YOR066W, as seen in the Y2H method, bound to protein kinase regulators, in the case of Clb2 and Clb5. In the Y2H method, YOR066W bound to a kinase, which phosphorylates amino acids, in the case of Cdc28. Mike Snyder's kinase functionality experiment would be a way to test the function of YOR066W. First, it would be necessary to make microwells to which different substrates would be covalently linked. One substrate would occupy each well. The YOR066W protein would then be added to each well along with radioactive ATP. After incubation, the microwell arrays would be imaged with a phosphoimager to see where radioactive Phosphates where incorporated into the substrates in the silicon. If YOR066W binds to kinases, this would suggest a function as a kinase substrate in the yeast genome. Experiment 2: Another possibile method to determine proteomic information about YOR066W is the isotope-coded affinity tags (ICAT) method. Since we are working with yeast, this experiment is not limited by the conditions of growing cells in heavy isotope. First, it is necessary to pick two conditions that will form different populations of cells. We could incubate one set of yeast cells in normal growth conditions and use the heavy ICAT labeling reagents to label these cells. Another set of yeast cells could be grown in a chemical condition that arrests the cell cycle at a particular phase. The cysteines of this second group of cells would be labeled with the light ICAT labeling reagent. After being labeled, the two cell populations would be mixed and biotin/avidin-purified. The labeled peptides would be put into a mass spectrometer to develop heavy to light protein ratio. Provided the protein ratios are big enough for the mass spec. to detect, we will see whether the cells that were arrested in a specific cell cycle produced fewer copies of any proteins. Of particular interest would be the affects of an arrested cell cycle on the YOR066W protein for which ratios could be compared to see if YOR066W plays a role in cell cycle. |
References
Campbell MA, Heyer LJ. 2003. Discovering genomics, proteomics, and bioinformatics. Bengamin Cummings: New York.
DIP. Database of Interacting Proteins. 2004 Nov 15 <http://dip.doe-mbi.ucla.edu/dip/Search.cgi?SM=3> Accessed 2004 Nov 17.
KEGG. Kyoto Encyclopedia of Genes and Genomes. 2004 May 31 <http://www.genome.jp/kegg/pathway/map/map01100.html> Accessed 2004 Nov 18.
MIPS. Munich Information Center For Protein Sequences. 2003 Nov 3 <http://mips.gsf.de/genre/proj/yeast/index.jsp> Accessed 2004 Nov 17.
PDB. Protein Data Bank. 2004 Nov 16 <http://www.rcsb.org/pdb/index.html> Accessed 2004 Nov 17.
SGD. Saccharomyces Genome Database. <http://www.yeastgenome.org> Accessed 2004 Nov 18.
YRC. Yeast Resource Center Two-Hybrid Analysis. 2000 July 11<http://depts.washington.edu/%7Eyeastrc/th_12.htm> Accessed 2004 Nov 17.
Emily Wilson's Genomics Home Page
Email Questions, Comments or Suggestions : emwilson@davidson.edu