This web page was produced as an assignment for an undergraduate course at Davidson College.
My Favorite Yeast Expression: SAE2 and YGL176C
In my last assignment, I examined two Sacchromyces cerevisiae genes, SAE2 and YGL176C, in depth using various databases. These two genes are juxtaposed on yeast chromosome 7. However, while SAE2 has been fully annotated, YGL176C has not yet been fully characterized. On this page, I will use expression data from the Expression Connection database to further analyze these genes.
Annotated Gene: SAE2
Summary from last time: SAE2 is a non-essential gene implicated in DNA double-strand break repair. SAE2 is transcribed in the cytoplasm and nucleus of cells.
Expression Data
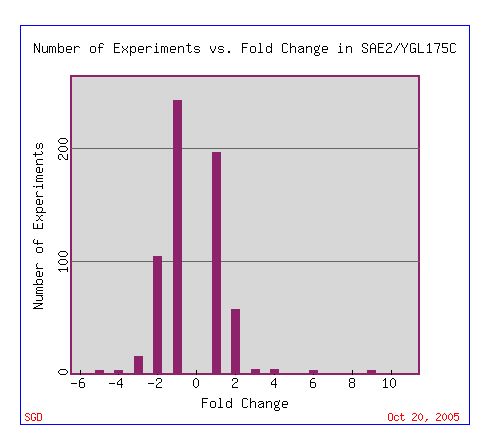
SAE2 mRNA expression has been examined in several experiments. Under certain conditions, SAE2 mRNA expression is either induced or repressed. The majority of the fold changes observed in SAE2 mRNA expression are small, ranging from -2 to 2 (Fig. 1).
Figure 1. Histogram depicting the number of experiments vs. the fold change in SAE2 mRNA expression. Positive numbers represent conditions under which SAE2 was induced, while negative numbers represent conditions under which SAE2 is repressed. Image courtesy of Expression Connection with a search for SAE2.
Minimal Induction and/or Repression of expression
Evident by Figure 1 above, most experiments demonstrated a minimal induction and/or repression of SAE2 mRNA expression. As just one pertinent example, Gasch AP, et al. (2001) examined SAE2 mRNA expression in wild type yeast and mutant yeast lacking mec1 when exposed to the DNA-damaging agents, methylating-agent methylmethane sulfonate (MMS) and ionizing radiation (Fig 2). Mec1 encodes a DNA- damage signal transducer.
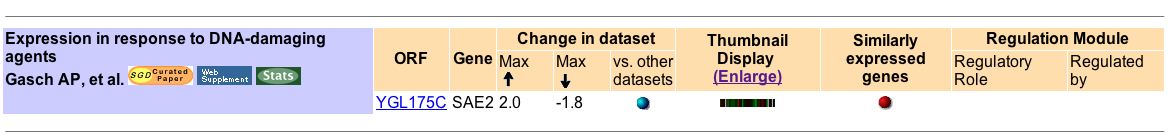
Figure 2. Summary of findings about SAE2 mRNA expression under DNA-damaging conditions. Image courtesy of Expression Connection with a search for SAE2.
They determined that under some DNA damaging conditions, SAE2 expression is induced, while under other DNA damaging conditions, SAE2 expression was repressed (Fig 3). For example, when wild type yeast are exposed to gamma irradiation, SAE2 is highly expressed at 5 minutes (Fig 3). Gamma irradiation breaks double-stranded DNA and SAE2 expression must be induced to produce SAE2p to repair the damage. At 10 to 30 minutes of being exposed to gamma irradiation, the wild type yeast show minimal or no induction of expression (Fig 3). Apparently more SAE2p is only good to a point and induction of SAE2 expression is not beneficial as DNA double-stranded breaks increase. Finally, SAE2 is repressed in wild type yeast when exposed to gamma irradiation for 45 to 60 minutes (Fig 3). SAE2p may actually be detrimental in yeast exposed to excessive levels of irradiation. So many DNA- breaks occur in the wild type yeast exposed to excessive gamma irradiation that the yeast undergo other repair pathways or apoptosis. The results of other experiments appearing below can be analyzed in a similar manner.

Figure 3. Microarray data for SAE2 expression in wild type and mec1 mutants when exposed to two different DNA-damaging agents. Red indicates induction of expression, green indicates repression of expression, black indicates neither induction nor repression of expression, and white indicates an error array at that spot/experiment. Image courtesy of Expression Connection.
No genes appear to be expressed in a similar manner in wild type or mec1 mutant yeast under these specific conditions.
Extreme Induction and/or Repression of expression
On the other hand, in rare experiments, SAE2 expression is extremely induced or repressed. In fact, Wyrick JJ, et al. (1999) found that SAE2 is generally highly induced or highly repressed under histone depletion. Histone is a protein essential to DNA supercoiling. Histone also plays a role in gene expression (i.e. genes that are supercoiled are not readily expressed) (Fig 4).
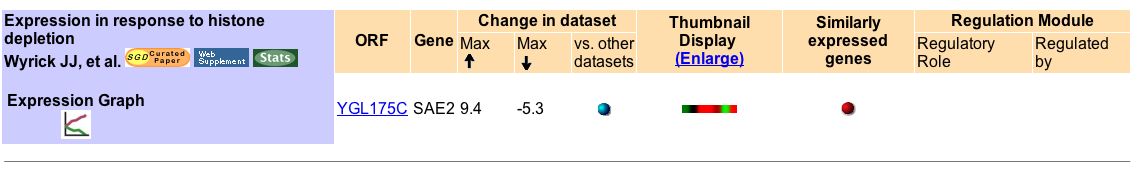
Figure 4. Summary of findings about SAE2 mRNA expression under histone depletion conditions. Image courtesy of Expression Connection with a search for SAE2.
The microarray data revealed a definite wave of repression to induction to repression to induction (Fig 5 and Fig 6). At zero hours of histone depletion, SAE2 expression is repressed (Fig 5). Thus, under normal condition, SAE2 is probably repressed. SAE2 is usually not necessary to repair double-stranded DNA breaks. There is no induction or induction of expression at 0.5 hours of histone depletion (Fig 5). At 1-2 hours of histone depletion, SAE2 expression is induced (Fig 5). Perhaps, since histone is depletion a wide range of gene normally repressed are induced because they are no longer supercoiled. Then, for some strange reason, at 4 hours SAE2 expression is repressed (Fig 5). Finally, at 6 hours SAE2 expression is highly induced again (Fig 5). It is unclear why a gene encoding for a DNA double-strand break repair protein would be strongly repressed after being strongly induced and then strongly induced a short while later.

Figure 5. Microarray data for SAE2 expression in yeast when histone is depleted. Red indicates induction of expression, green indicates repression of expression, and black indicates neither induction nor repression of expression. Image courtesy of Expression Connection.
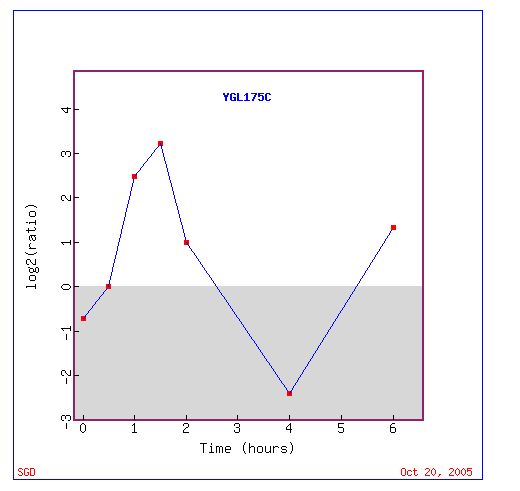
Figure 6. Graphical representation of the shift over time from repression to induction to repression to induction of SAE2 during histone depletion. Log2(ratio) indicates the Log2(red/green). Negative values of Log2(ration) indicate higher amount of green (repression), while positive values indicate higher detection of red (induction). Image courtesy of Expression connection.
There are several genes that exhibit a similar expression pattern under histone depletion over time (Fig 7). Some annotated genes have a similar role to SAE2, like MRE11, which is also involved in DNA repair. Other annotated genes that cluster with SAE2 based on expression data do not appear to have a similar processes, functions, or components, like NGG1 and FLO8. Perhaps these genes simply react similarly to histone depletion conditions and are not otherwise related to SAE2. Thus, the non-annotated genes appearing below, like YMR041C, may or may not have similar function to SAE2.
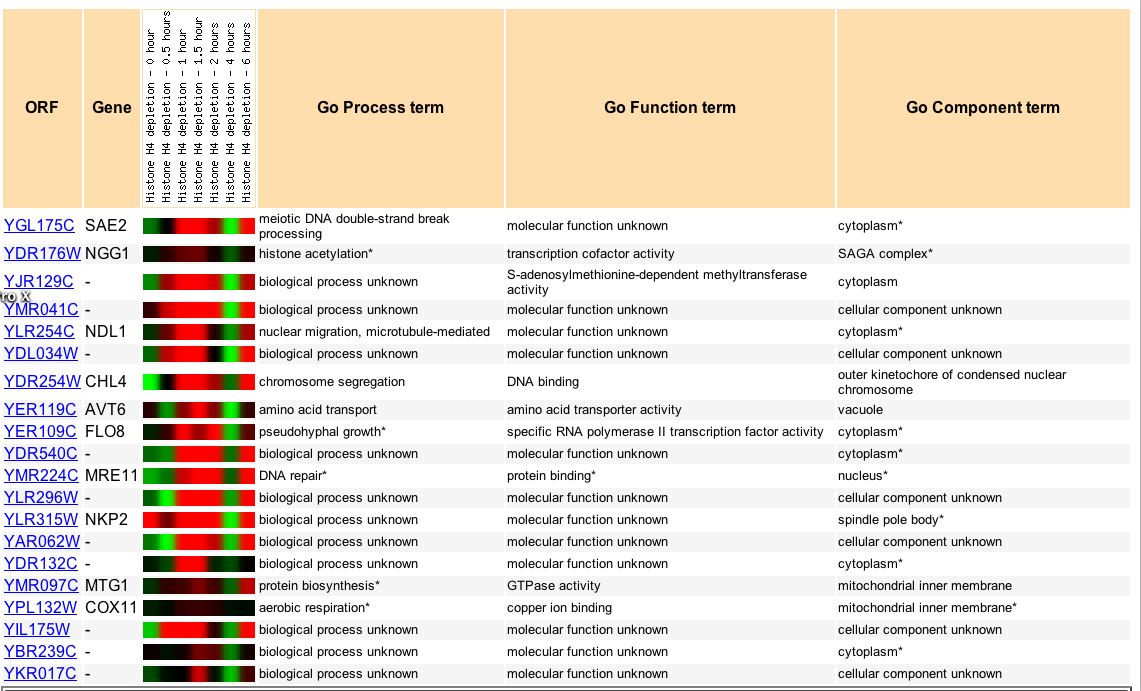
Figure 7. Genes that have similar expression patterns to SAE2 under histone depletion conditions. Image courtesy of Expression Connection.
Non-Annotated Gene: YGL176C
Summary from last time: YGL176C is a non-essential gene that currently has no known molecular function, biological process, or cellular component. I hypothesized that the protein product of YGL176C may be a receptor and/or that it may take part in folding proteins.
We can use microarray expression data to develop a better hypothesis as to the function of YGL176C. As evident by the histograph below, many experiments have been conducted that measure how various conditions affect the expression pattern of YGL176C mRNA. Most experiments measured only minor fold changes, but some experiments showed extreme fold changes in the expression of YGL176C mRNA (Fig 8).
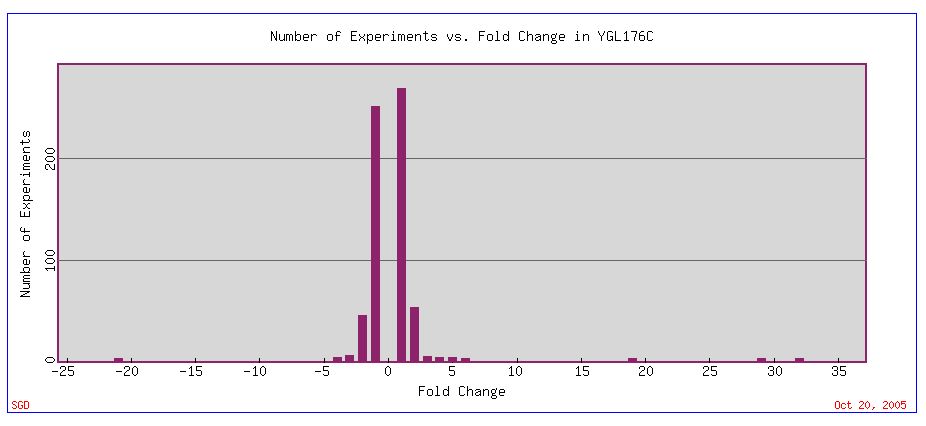
Figure 8. Histogram depicting the number of experiments vs. the fold change in YGL176C mRNA expression. Positive numbers represent conditions under which YGL176C was induced, while negative numbers represent conditions under which YGL176C is repressed. Image courtesy of Expression Connection with a search for YGL176C.
In fact, Derisi JL, et al (1997) investigated the change in yeast gene expression as the yeast shifted from the anaerobic process of fermentation to aerobic respiration (diauxic shift). They found that YGL176C expression is repressed at the beginning of diauxic shift (Fig. 9). Eventually, however, YGL176C expression is induced (Fig. 9). Therefore, perhaps YGL176C is normally expressed under aerobic conditions and repressed under anaerobic conditions. Maybe YGL176C is not useful until the late stages of diauxic shift.
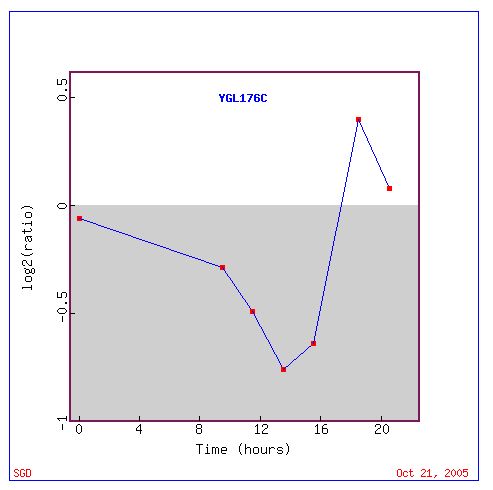
Figure 9. Graphical representation of the shift over time from repression to induction of YGL176C during diauxic shift. Log2(ratio) indicates the Log2(red/green). Negative values of Log2(ration) indicate higher amount of green (repression), while positive values indicate higher detection of red (induction). Image courtesy of Expression connection.
Analyzing how YGL176C expression changes under diauxic shift is most useful when compared to the expression patterns other genes of known function also under diauxic shift. We can search for genes that show a similar expression pattern to YGL176C. We can use guilt by association to hypothesize how YGL176C might function. Under diauxic conditions, YGL176C clusters with a wide variety of genes (Fig 10). It is difficult to see a pattern in the genes with which YGL176C clusters because many of the genes also have no known process or function.
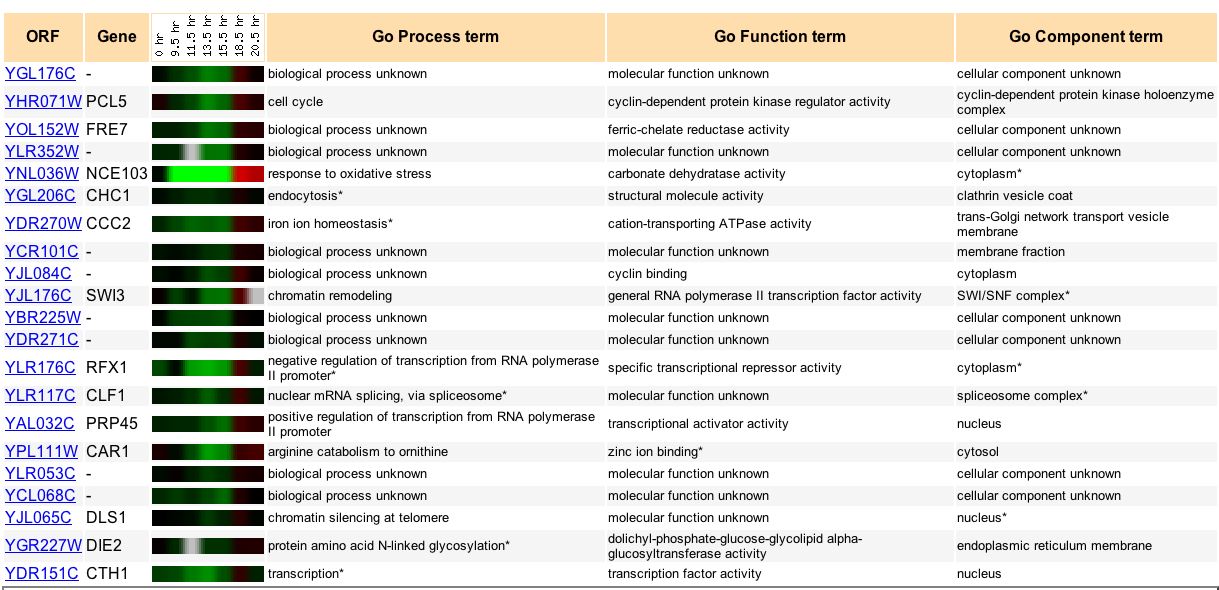
Figure 10. Genes that have similar expression patterns to YGL176C under diauxic shift. Image courtesy of Expression Connection.
We can perform a gene ontology term search on the SGD website to better analyze similarity in the genes with which YGL176C clusters under diauxic shift. As evident in the figure below, many of the genes with which YGL176C clusters are involved in maintaining homeostasis (Fig. 11). Perhaps YGL176C is involved with homeostasis, too.
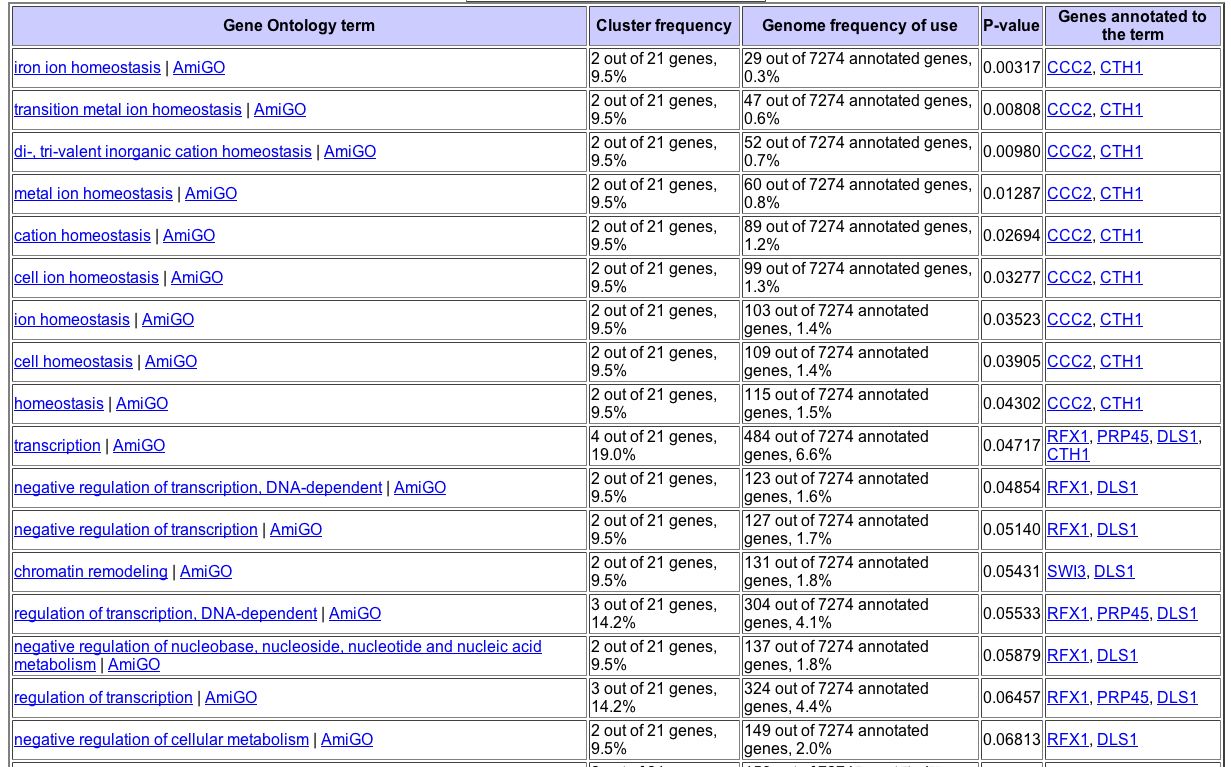
Figure 11. Onotology of genes that cluster with YGL176C based on diauxic shift. Image courtesy of SGD Gene Ontology term finder, with a search for gene that had a similar expression pattern to YGL176C during diauxic shift.
We can also examine how YGL176C expression relates to genes under other conditions. Travers KJ, et al. (2000) exposed yeast to endoplasmic reticulum stress and measured the unfolded protein response. Surprisingly, they found that "rather than regulating only ER-resident chaperones and phosphor biosynthesis, as anticipated from earlier work, unfolded protein resonse affect multiple ER and secretory pathway functions" (Travers KJ, et al., 2000). In wild type yeast exposed to endoplasmic reticulum stress, YGL176C expression is highly induced (Fig. 12). Based on its expression pattern under ER stress, YGL176C clusters with several genes of known function (Fig. 12).
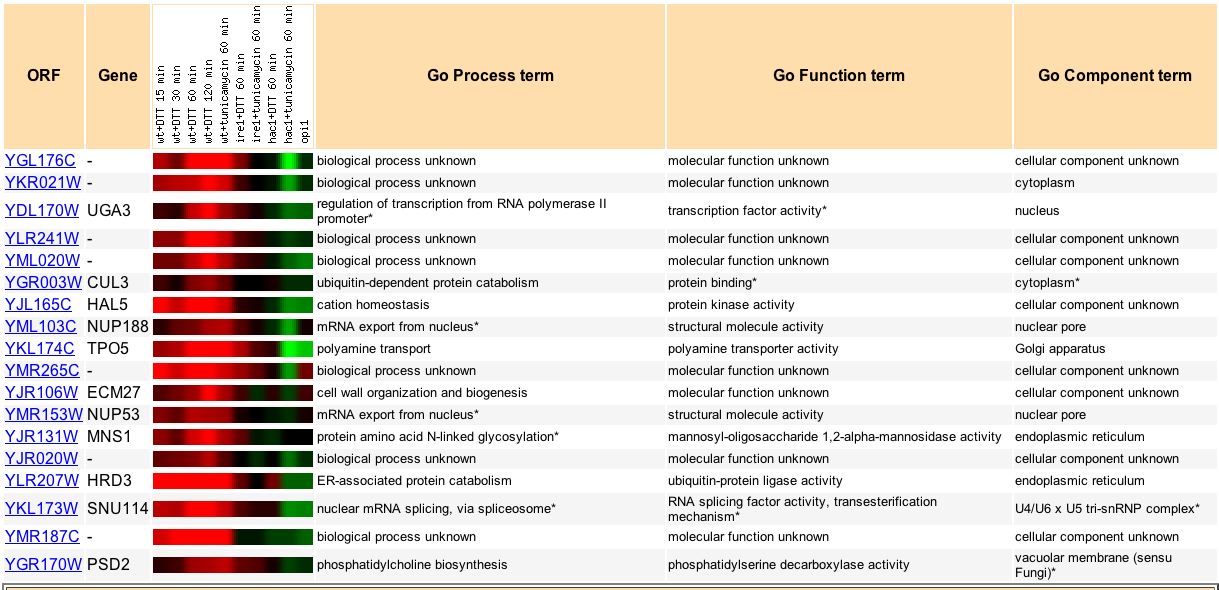
Figure 12. Genes that have similar expression patterns to YGL176C during unfolded protein response. Image courtesy of Expression Connection.
By using a computer to compare the functions of all genes that clustered with YGL176C during ER stress, a distinct pattern emerges. Many of the genes that have a similar expression pattern to YGL176C under ER stress are involved in transport of some form or another (Fig. 13). Therefore, using guilt by association, YGL176C may have a role in transport of some sort.
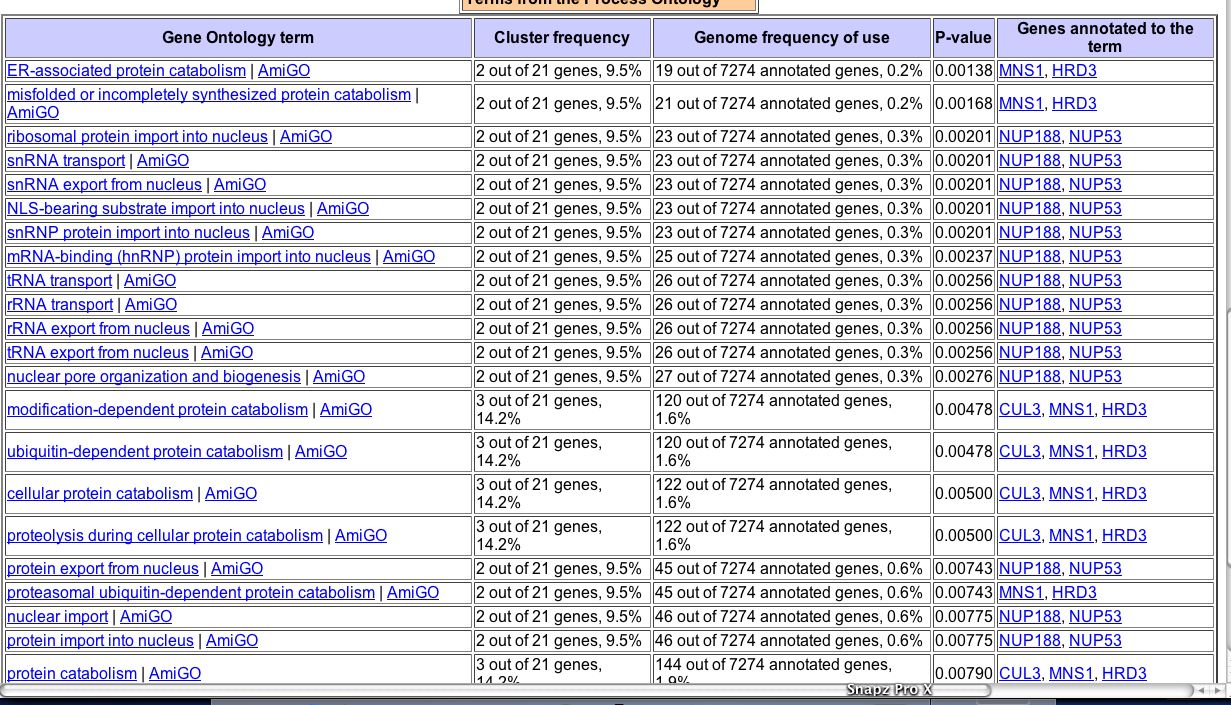
Figure 13. Genes that cluster with YGL176C during the stress to the ER. Cluster Image courtesy of SGD Gene Ontology term finder, with a search for gene that had a similar expression pattern to YGL176C during unfolded protein response.
Finally, Ferea TL, et al. (1999) examined the expression pattern of various yeast genes when the yeast were grown for many generations under glucose-limiting conditions. They found that yeast strains evolved with alteration of expression of genes "involved in glycolysis, the tricarboxylic acid cycle, oxidative phsophorylation, and metabolite transport" (Ferea TL, et al., 1999). Specifically, Ferea TL, et al. (1999) found that evolved 2 and 3 strains of yeast had repressed expression of YGL176C compared to the parent. Perhaps the expression of YGL176C is not beneficial or even harmful to energy deprived yeast. YGL176C is probably not involved directly in glycolysis, the tricarboxylic acid cycle, or oxidative phosporylation. If YGL176C is involved in transport it is probably uses a lot of energy and would logically be repressed under energy limiting conditions.
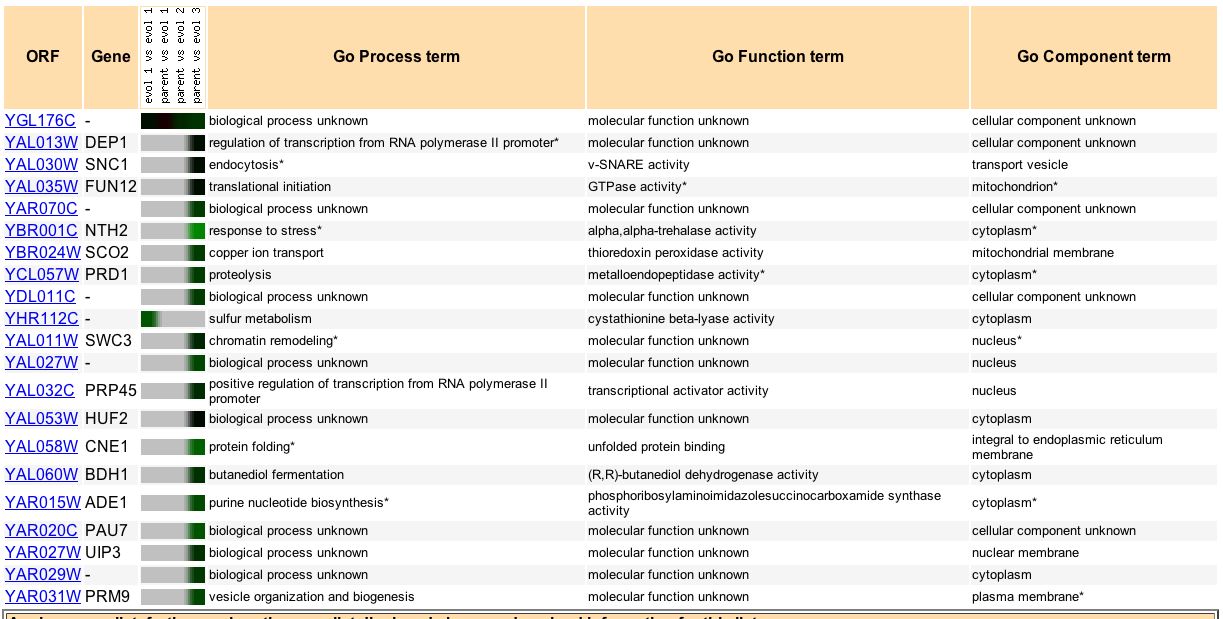
Figure 14. Genes that have similar expression patterns to YGL176C during glucose limiting conditions. Image courtesy of Expression Connection.
The genes with which YGL176C clusters during glucose-limiting conditions are not particularly helpful in narrowing down the function of YGL176C. Only a few of the genes with which YGL176C clusters have a known function and of those only two similar ontologies emerge (Fig. 14 and Fig. 15). In this case, guilt by association would predict that YGL176C may be involved with proteolysis and/or the regulation of transcription from RNA polymerase II promoter I (Fig. 15).

Figure 15. Glucose Limitation Cluster. Image courtesy of SGD Gene Ontology term finder, with a search for gene that had a similar expression pattern to YGL176C under glucose limiting stress.
Revised Hypothesis regarding YGL176C
Using all of the expression data above, I have a revised hypothesis about the role of YGL176C. I initially predicted that this gene encoded a protein that was receptor and/or involved in protein folding. In light of the expression data , instead proposed that YGL176C is a protein involved in maintaining cellular homeostasis by transporting molecule across membranes. A receptor and a transmembrane protein transport molecule have similarities in that both have transmembrane regions and an abundance of alpha-helices. Likewise, often both require energy to function. It is important to note that nevertheless, YGL176C may play a role in protein folding or possess some totally different function. The microarray data do not overwhelmingly support these alternative functions, however.
References:
Balakrishnan R, Christie KR, Costanzo MC, Dolinski K, Dwight SS, Engel SR, Fisk DG, Hirschman JE, Hong EL, Nash R, Oughtred R, Skrzypek M, Theesfeld CL, Binkley G, Lane C, Schroeder M, Sethuraman A, Dong S, Weng S, Miyasato S, Andrada R, Botstein D, Cherry JM. 2005. Saccharomyces Genome Database. < http://www.yeastgenome.org/>. Accessed 2005 Oct 20.
DeRisi JL, Iyer VR, Brown PO. 1997. Exploring the metabolic and genetic control of gene expression on a genomic scale. Science 278: 680-686
Ferea TL, Botstein D, Brown PO, Rosenzweig RF. 1999. Systematic changes in gene expression patterns following adaptive evolution in yeast. Proc Natl Acad Sci U S A 96: 9721-9726
Gasch AP, Huang M, Metzner S, Botstein D, Elledge SJ, Brown PO. 2001. Genomic expression responses to DNA-damaging agents and the regulatory role of the yeast ATR homolog Mec1p. Mol Biol Cell 12: 2987-3003.
Travers KJ, Patil CK, Wodicka L, Lockhart DJ, Weissman JS, Walter P. 2000. Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell 101: 249-258.
Wyrick JJ, Holstege FC, Jennings EG, Causton HC, Shore D, Grunstein M, Lander ES, Young RA. 1999. Chromosomal landscape of nucleosome-dependent gene expression and silencing in yeast. Nature 402:418-423.
Return to Jackie's Genomics Main Page