Evolutionarily Assembled cis-Regulatory Module at a Human Ciliopathy Locus
This web page was produced as an assignment for an undergraduate course at Davidson College.
This webpage summarizes the article "Evolutionarily Assembled cis-Regulatory Module at a Human Ciliopathy Locus" authored by Lee et al. and explains the figures presented within the article.
Preliminary Thoughts
I am appreciative and fascinated by the work done in this article because it test the boundaries of nonparalogous genes as we know it in addition to discovering another evolutionary aspect of human (and other higher order
vertebrates) development. Not only did the experiementers conduct a thorough study, they presented the material in a way that is reader friendly and informative.
Information to Note
- Little is known about nonparalogous gene clusters; however, scientists have observed their existence.
- Evidence has shown that cilia play very important roles in vertebrate development. For example, primary cilia can serve as cellular antennae and hence be able to detect a variety of signals.
Introduction
One of the most common neurodevelopmental disorders associated with the ciliopathy system is Joubert Syndrome (JBTS). Symptoms of the disorder could include abnormal eye movement, hypotonia, and an obvious mid-hindbrain malformation. Researchers have been trying to find causes for this disorder and results have shown that in half of the patients, of a 10 patient sample; there was a mutation on the transmembrane protein (TMEM) 216 gene, which was linked to the JBTS2 locus on chromosome 11. The other half of the patient sample did not have any indication of the mutated TMEM216 gene; however, they possessed the same phenotypic expression as the first half. This lead researchers to conclude that there was another causative gene operating at the JBTS2 locus.
Researchers then decided to resequence all the known and predicted exonic and genetic promoter elements, hence discovering four missense mutations and one splicing homozygous deleterious mutation in conserved areas near the TMEM138 gene, which has no known function. Even though the gene has no known function, it is known that mutations in the TMEM138 p.H96R transmembrane domain lead to unstable protein, potentially resulting in the loss of function as the disease mechanism. These mutations were found in all of the JBTS2- linked families.
This was initially puzzling for researchers because although the two TMEM genes encoded transmembrane proteins, they were not shown to be homologs or have any shared functional domains. They were considered distinct protein families; however, this view will soon change. The challenge was now to figure out how these two genes relate to each other in order to figure out how they co-function in JBTS.
The Challenge
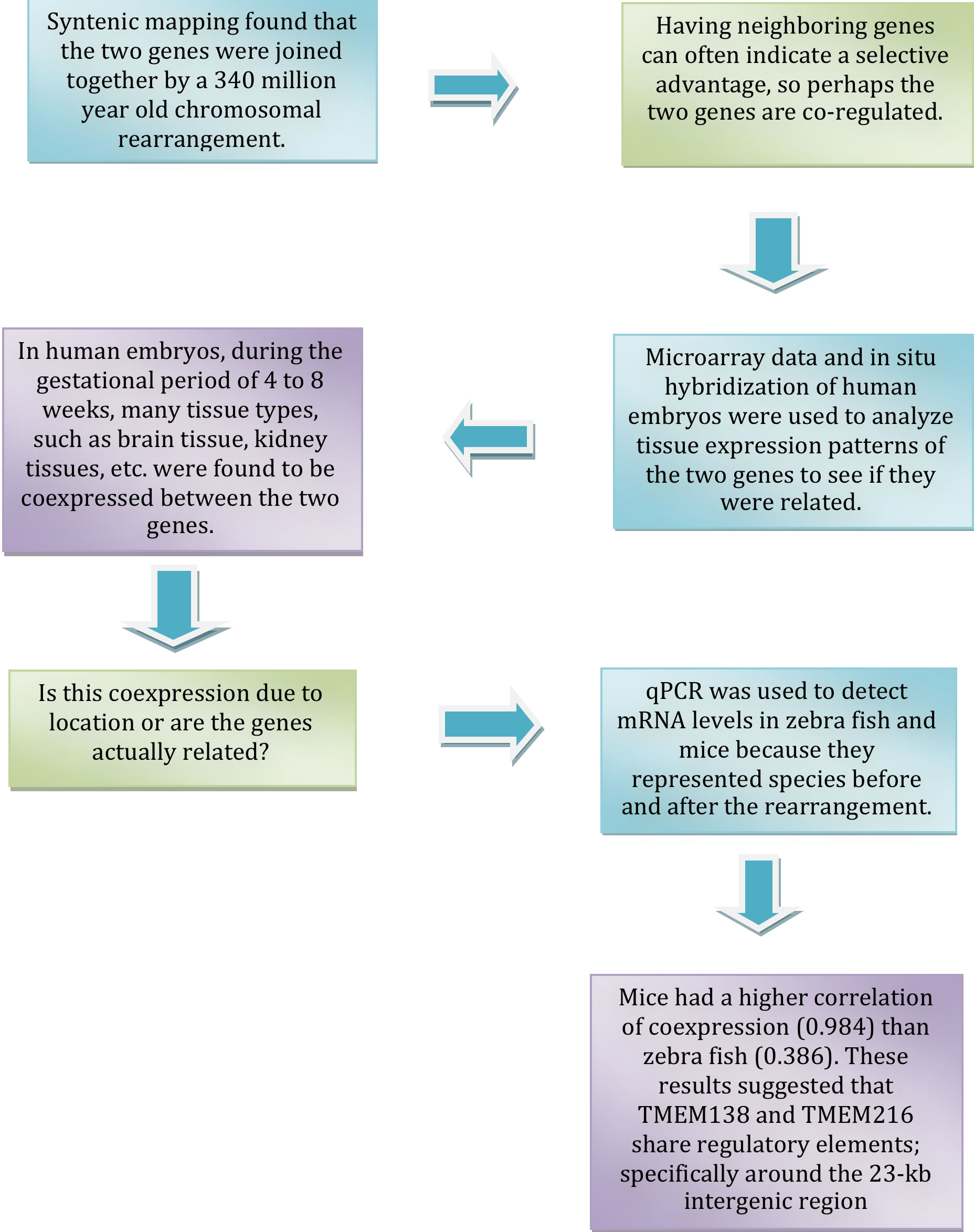
Researchers conducted the following main test to identify links between the two genes. The process is as follows: (Blue boxes indicate the tests conducted. Green boxes indicate the thought process behind the conducting the test. Purple boxes indicate results.)
Conclusions
Other minor tests were conducted; however, from the culmination of results, researchers were left to conclude that the two genes did indeed exhibit coordinated expression. This finding, coupled with the fact they produced similar phenotype, suggested that the gene could also be relate on the protein level. This assumption was validated and expounded upon by another experiment, whose results showed that ciliogenesis, the production of cilia, became defective after a knockdown of TMEM138 and TMEM216 in IMCD3 cells. Hence demonstrating that not only are the coexpressed; they also play a role in the production of cilia.
Further testing geared towards discovering the genes’ mechanistic role in ciliogenesis was conducted and results found that TMEM138 and TMEM216 were located near linked but distinct vesicle pool which indicated that they direct en route vesicles to the base of the cilium, which is important for ciliary assembly. It was also shown that the genes could have a dependency on one another for certain aspects of vesicular movement.
The last and most important finding that scientists uncovered was that before the genes were rearranged to be proximal to each other, their protein functions were not the same and their phenotypic expression was different as well. This is an interesting finding because it demonstrated that their change in location, relative to each other, was a determining factor of their linked behavior expressed in the present. In other words their coordinated expression was not built in but evolved. This case provided an example that nonparalogous genes could be rearranged to form functional gene clusters by evolution.
Breaking Down The Figures and Tables
A. This figure shows the location of the TMEM138 and TMEM216 genes (indicated in red letters). They are located on chromosome 11 in the JBTS2 locus in the p.11.2-q13.1 position in a head-to-tail configuration, with TMEM138 being located 23 kb away from the tail of TMEM216. The diagram goes on to magnify the genes to show more intricate detailing of the genes. The TMEM138 gene encodes a tripsan membrane protein and the TMEM216 gene encodes a terapan membrane protein. It is worth noting that the TMEM138 gene has a signal peptide; whereas the TMEM216 gene does not. Both have alternating transmembrane and extratransmembrane regions.
B. The diagram shows the syntenic mapping done for the TMEM138 and TMWM216 genes. The different species are all related and are ordered such that the oldest species (before the gene rearrangement) is at the bottom and the newer species (after the gene rearrangement) are at the top. The different colors represent different conserved (intergenic regions, exons, introns, and UTRs) and the black represent the unconserved intergenic, exonic, and intronic of the chromosome. One can see the progression from the zebra fish having the genes on two separate chromosomes to the anolis which has the genes on the same chromosome but has unconserved regions. Slowly one can see (beginning with the mouse) the progression of the species accumulating more and more conserved regions. Even though the lower species have some conserved exons, they do not possess any other conserved regions. The take home message from the map is that the conserved regions on the chromosome are can be found in species from reptiles to humans; however, they are not present in lower vertebrates.
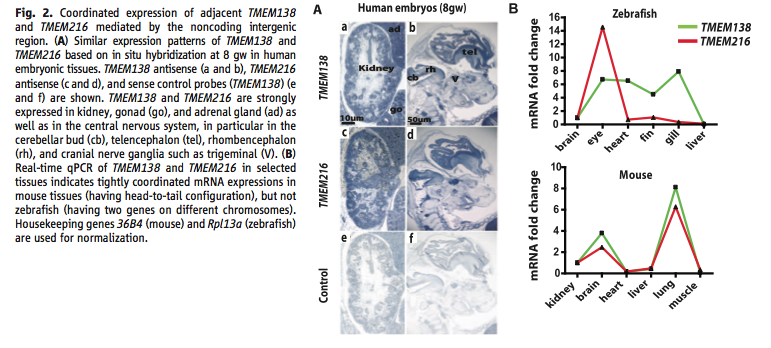
A. The illustrations depicts the results from the in situ hybridization experiment that tested for coordinated expression between the two genes in human embryonic tissues. The rows indicated the experimental environment that produced the sample (TMEM138 antisense, TMEM216 antisense, and the sense control). As one could see the, by the colored shading, that both of the genes have very similar expressions especially in the kidneys, gonad(go) and adrenal (ad) glands(shown in a) and in the cerebellar bud (cb), telencephalon (tel), rhombencephalon (rh), and cranial nerve ganglia (V). The control condition showed a different expression than the genes.
B. The graphs represent the mRNA expression of TMEM138 (green lines) and TMEM216 (red lines) via mRNA fold change in zebra fish and mice. Zebra fish represent species occurring before the gene rearrangement (having the two genes on different chromosomes) and mice represent species occurring after the rearrangement (the genes being in a head-to-tail conformation). The graphs compare the mRNA fold change in different bodily tissues. Demonstrated by the closeness or lack of closeness of the lines to each other, one can detect how tightly coordinated the mRNA expressions were. Mice had a higher correlation (0.984) between the coordination of the two genes than zebra fish (0.386), indicating that the head-to-tail conformation of the genes is important for their co-regulation.
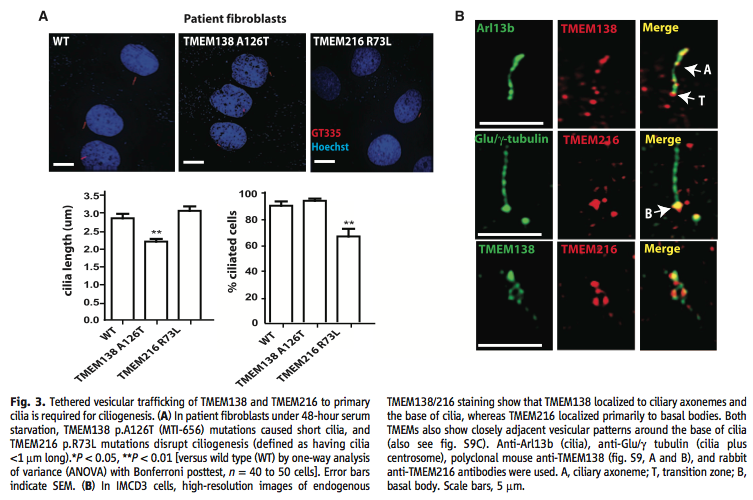
A. The figure shows the results from the experiment that tested whether or not the genes demonstrated a functional relationship at the protein level. The illustrations above the graph show patients fibroblast that were deprived of serum for 48hours. The wild type served as a comparison in pictorially showing the length of the cilia. It was shown that mutations in TMEM138 p.A126T resulted in shorter ciliaa lengths (shown as statically significant in the following graph) and mutations in TMEM216 p. R73L resulted in a disruption of ciliogenesis (shown as having a significantly lower percentage of ciliated cells). The graphs below quantify the actual length of the cilia in the three conditions and then provide a percentage of how many cells actually grew cilia. The graphs validates the aforementioned conclusions.
B. The illustration shows the results from the experiment in which researches discovered that TMEM216 was mainly located near the basal bodies; whereas TMEM138 was located near ciliary axonemes and the base of the cilia. Both of the genes showed close vesicular patterns around the base of the cilia. The row represent the different cilia conformations analyzed. Arl13b shows a cilia, Glu/-?tubulin shows a cilia plus the centrosome, and anti-TMEM138 antibodies from polyclonal mice and anti-TMEM216 antibodies form rabbits.
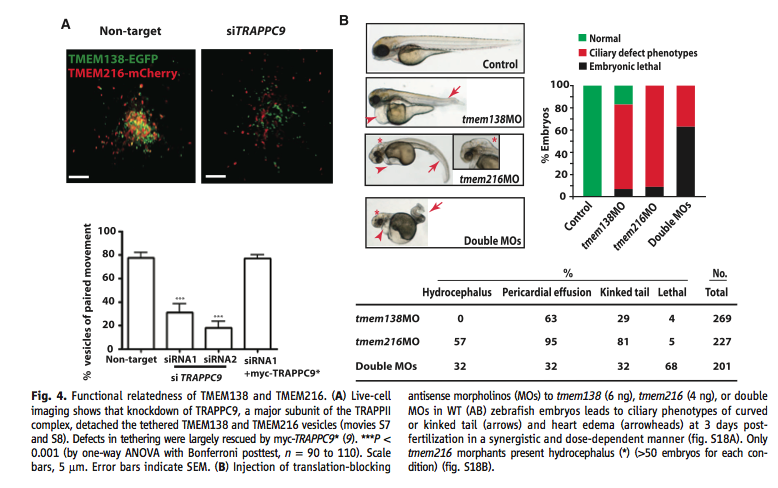
A. The cell imaging illustration and graph below shows the importance of the TRAPPII (tethering) protein by knocking down TRAPPC9 which is a major subunit of TRAPPII. TRAPPII mediates the tethering of the Golgi vesicles in TMEM138 and TMEM216 that are important for ciliary assembly. In tethering, vesicle links are spread over a distance that is more than half the size of the vesicle's diameter from its corresponding membrane, hence making vesicle binding closer to each other. In the knockdown condition (siTRAPPC9) one can see that the vesicles are more spread out than in the control condition. The graph shows that in the conditions that were experienced the TRAPPC9 knockdown had significantly less vesicles of paired movement.
B. This figure demonstrates the effects that translation-blocking antisense morpholinos (MOs) have on zebra fish embryos. Each gene had their own injected MO. When the zebra fish embryo was injected with the TMEM138 MO the embryos showed a heart edema and a slight kin in the tail (shown by the arrows). When injected with the TMEM216 MO, the embryos showed a significantly greater amount of hydrocephalus and a severely kinked tail (shown by the arrows and the asterisk). When both of the MOs were injected, the embryos showed a disfigured tail, hydrocephalus, and an enhanced heart edema(shown by the arrows and the asterisk). The graph shows the percentage of embryos that were normal, defected, or died depending of which MOs were injected. In the control 100% of the embryos were normal which indicates that the sample had healthy embryos. In the TMEM138 MO condition, there was a small percentage of embryos that developed; the majority of them had defective phenotypes, and a relatively smaller percentage of them died. However, in the TMEM216 MO condition, there were no normally developing embryos; the majority had defective phenotypes, and a small (but slightly greater than in the TMEM138 MO) died. In the double dosage condition, the majority of the embryos died and the remaining percentage had defective phenotypes. The chart shows that percentage of embryos in the manipulated conditions that demonstrated hydrocephalus, pericardial effusion, a kinked tail, and that died.
Final Thoughts
The work conducted in this article is revolutionary becasue it demonstrates the power of evolution in that higher level vertebrates are able to have nonparalogous gene that are coexpressed. Not only are the experiments
groundbeaking in that they are able to produce such meaningful results in seemingly simple ways, the actual findings from these results have reshaped the way organisms are seen. Species are constantly evolving and everyday
scientist are uncoering new tidbits that shed light to the complex inner working of the complex organisms.
References
Daghlian, C. (Photographer). (n.d.). Lung trachea 1a. [Print Photo]. Retrieved from http://remf.dartmouth.edu/images/mammalianLungSEM/source/9.html
Lee, J. et al., (2012, February 24). Evolutionarily assembled cis-regulatory module at a human ciliopathy locus. Science, 335(6071), 966-969. Retrieved from http://www.sciencemag.org/content/335/6071/966.full?sid=cf3e3faa-8 170-4eae-9b5c-78d55356053f
Genomics Page
Biology Home Page
© Copyright 2012 Department of Biology, Davidson College, Davidson, NC 28035