This web page was produced as an assignment for an undergraduate
course at Davidson College.
Single Molecule Protein Interactions
What do proteins do?
Proteins are central to the fundamental chemistry of life. Cells
are a buzzing milieu of diverse proteins performing a wide range of
functions. Proteins cooperate to execute many complex tasks, such as
transmitting information along signaling pathways and assembling and
disassembling molecules during metabolism.
Where is the protein located?
Our current understanding of fundamental cellular processes has emerged
from a combination of methods, including direct observation and
indirect experiments. Direct observation is appealing, because it
enables researchers to see how proteins fit within the spatial context
of the cell. Viewing the locations of proteins in a live cell often
relies on the small protein GFP, which remarkably fluoresces green
without any need for added substrates. By genetically fusing GFP to a
protein of interest, we can identify the subcellular localization of
the protein using fluorescence microscopy. Portions of the cell
containing the protein fluoresce green.
In most applications, the resolution available to conventional
fluorescence microscopes is insufficient to track single proteins
(reviewed in Huang et al., 2010). Inevitable slight imprecisions in
focus are problematic at high magnification. Despite the limitation,
GFP tags provide excellent information about the general location of
proteins within the cell.
What is the protein’s structure?
We can, however, resolve the structure of a protein in atomic detail
using X-ray crystallography. Growing protein crystals is challenging,
and success requires good fortune as much as it does technical
expertise, but the results are stunning. The structure of GFP (see the
image) reveals a barrel shape made of beta sheets, called a “beta
barrel.” Inside the beta barrel is the chromophore, responsible for the
fluorescence that makes GFP useful to researchers.
Crystal structures can both test and generate hypotheses about the
activities performed by the many proteins in a cell. Additional
experiments are required to interrogate proteins in their native
physiological contexts, such as how often another protein is bound and
what consequences result from the binding event.
 Green fluorescent protein, GFP
Green fluorescent protein, GFP.
The chromophore, responsible for the fluorescence mechanism of GFP, is
colored green and not to scale. Coordinate data obtained from PDB ID
1EMA and rendered with Jmol.
What interacts with the protein?
Proteins do not act in isolation. Interactions between proteins
are key to many biological processes. Immunoprecipitation and Western
blotting have been used to map out many biological pathways where
protein-protein interactions serve an important role. Western blots use
an antibody against a protein of interest to test for its presence in a
sample, and immunoprecipitation generates samples containing the
binding partners of a protein. Together, they can tell us if two
proteins interact in a particular organism, tissue, or physiological
state.
But, ultimately, Western blots are averages. Retrieving enough protein
to see a band in the blot requires pooling the lysates from many cells.
Through this averaging process, Western blots hide the biological
variation that exists from cell to cell, and from protein to protein
within each cell. The nuance between the two questions, 'Do proteins
interact?' and 'How do proteins interact?' is not trivial. Biological
systems are dynamic, complex, and sometimes chaotic. A mechanistic and
quantitative understanding requires precision in details.
How do proteins perform their functions?
Fluorescence
microscopy, X-ray crystallography, and immunoprecipitation can provide
fundamental information about what a protein does. However, studying
proteins in bulk can hide insights that arise from appreciating the
variation that exists or examining cells at a very small scale. In
contrast, through examining proteins individually, single molecule
experiments can reveal unexpected behaviors and the diversity that
exists in cells.
How does an enzyme interact with its substrate?
Cholesterol oxidase, as the name suggests, oxidizes cholesterol (Lu et
al., 1998). The net reaction is loss of electrons by cholesterol and
gain of electrons by oxygen, with hydrogen balancing the charges. The
reaction mechanism requires that the active site of cholesterol oxidase
bind the coenzyme FAD. Electrons are transferred from cholesterol to
FAD in the first step of the reaction, and subsequently passed along to
oxygen.
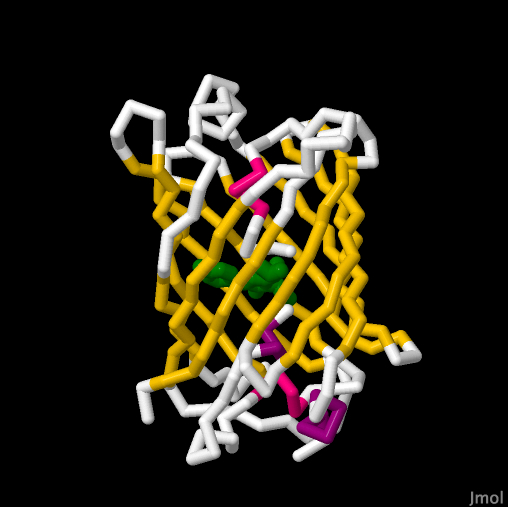
 Cholesterol oxidase
Cholesterol oxidase. The active
site of cholesterol oxidase binds the coenzyme FAD (highlighted in
light blue) to catalyze the oxidation of cholesterol. During the
reaction, FAD accepts electrons from cholesterol prior to transfer to
oxygen. In its electron-bound state, FAD cannot fluoresce. Coordinate
data obtained from PDB ID 1B4V and rendered with Jmol.
FAD is fluorescent, but in an electron-bound state its ability to
fluoresce is suppressed. Therefore when bound to cholesterol oxidase
you would expect each FAD molecule to transition back and forth between
fluorescent and non-fluorescent states, depending on the progress of
the reaction. In other works, FAD should blink on and off: on by
default, but switched off in the intermediate reaction state where
electrons have been taken from cholesterol but have not yet been handed
off to oxygen.
Flashes of FAD fluorescence are precisely what Lu et al. found. In
their experiment, the authors used purified cholesterol oxidase
embedded in a gel containing the substrate cholesterol. The surface of
the gel was illuminated with light to excite the FAD molecules, and a
camera detected any light emitted.
By combining data obtained from observing many individual enzymes in
the gel matrix, they found that the reaction followed unexpected
kinetics. The rates of FAD flashes did not fit the original prediction.
By generating a mathematical model and testing the model against their
experimental data and against computer simulations, they concluded that
the best explanation for the data was that reaction rate depended on
which of two major conformations the enzyme exhibited at a given time.
In other words, the efficiency of the enzyme depended on its ability to
change conformations between individual reactions. Once the reaction
was complete, the protein was not necessarily ready to accept more
cholesterol. A conformation change must occur first, a step ignored by
conventional models of enzyme kinetics. Proteins are more flexible than
static images tempt us to believe, and enzyme efficiency depends on the
individual history of each molecule.
Motor proteins: the ultimate single molecule experiments
Experiments like observing cholesterol oxidase can help us
understand how single enzymes behave, but no field has benefited more
strongly from single molecule experiments than study of the cell
biology of motor proteins.
The eukaryotic cytoskeleton not only provides structural rigidity. It
is also a dynamic transportation network for organelles, particles of
mRNAs bound to protein, and other cellular cargo. The motor proteins
dynein and kinesin walk (literally!) along microtubules. Members of the
dynein protein family walk in the opposite direction as those of the
kinesin family. While the proteins are quite different in structure,
they both contain two domains that can bind tubulin, the monomer used
to construct microtubules. Alternate binding of the two domains causes
the motor protein to walk or slide along the microtubule, dragging
attached cargo along with it.
If microtubules are organized like a highway system, we might expect
particular motor proteins to travel only along a subset of
microtubules. It would be much more efficient for a motor protein
destined for the cell membrane to travel along the most direct route
possible, rather than detouring halfway to visit the Golgi.
We know that the microtubules in a cell are not all the same; there are
subpopulations. But how these differences translate into specialized
transport of cargo has not been as clear. Through single molecule
experiments, Cai et al. (2009) found that the Kinesin-1 motor localizes
to a subset of microtubules (see the figure). In the image,
microtubules are labeled with the fluorescent protein mCherry (shown in
red), and Kinesin-1 by the fluorescent protein mCitrine (shown in
green). Rather than seeing spots of mCitrine-labeled Kinesin-1
scattered across the mCherry-labeled microtubules, they found that the
proteins are limited to a relatively small fraction of microtubules.
This suggests that Kinesin-1 does not bind to microtubules randomly,
but the experiment does not exclude the possibility of a burst of
activity, such as many vesicles being trafficked in a similar way at
one time.
An additional experiment clarified the ambiguity, showing that the
microtubules ‘preferred’ by Kinesin-1 are marked with
post-translational modifications, which is associated with stable
microtubules. It appears that Kinesin-1 is specialized to the 'tried
and true' part of the cytoskeleton, while other motor proteins walk
along parts of the cytoskeleton being actively assembled and
disassembled.
 High resolution microscopy of Kinesin-1 on microtubules.
High resolution microscopy of Kinesin-1 on microtubules. Kinesin-1
proteins are shown in green, and microtubules in red. Kinesin-1
localizes to a subpopulation of microtubules, rather than being broadly distributed across the cell. Scale bar indicates 4 µm.
Image obtained from Cai
et al. (2009), Figure 1G (reproduced under terms of the
CC BY license).
Conclusion
The question 'What does this protein do?' is complex and
multifaceted. Immunoprecipitation, Western blotting, and X-ray
crystallography are central to gathering fundamental information about
proteins, but they do not provide a complete story. Proteins are
autonomous machines, and experiments that look at proteins in aggregate
will inevitably miss crucial parts of the story. Single molecule
experiments reveal the underlying variation and unexpected behaviors
that would otherwise be lost to an average.
References:
Huang B, Babcock H, Zhuang X. 2010. Breaking the diffraction barrier: super-resolution imaging of cells. Cell 143:1047-1058.
Lu HP, Xun L, Xie XS. 1998. Single-molecule enzymatic dynamics. Science 282:1877-1882.
Cai
D, McEwen DP, Martens JR, Meyhofer E, Verhey KJ. 2009. Single molecule
imaging reveals differences in microtubule track selection between
kinesin motors. PLoS Biol 7:e1000216.
Eric Sawyer's Home Page
Genomics Page
Biology Home Page
Email Questions or Comments.
© Copyright 2014 Department of Biology, Davidson College,
Davidson, NC 28035