This web page was produced as an assignment for an undergraduate course at Davidson College.
Summary:
This article questions the idea of duplicated genes in yeast protecting the cells from genetic perturbations. It has been thought that gene duplication has been an important process throughout evolution, where the spare copy of the gene can acquire new functions. This article measured the interaction of yeast proteins with their partners in wild-type cells and in cells that lack a paralog. Approximately half of the paralog pairs compensate for the lost interactions and the other half maintain the interactions. The article provides data trying to argue that gene duplications might increase mutational fragility instead of robustness.
The research focused on 56 pairs of paralogous proteins with which protein-protein interactions (PPI) can be studied in vivo while looking at different genetic backgrounds. A PPI network of these 112 proteins was mapped in the wild type background and in the deletion background of their cognate copy. The researchers then performed a protein fragment complementation assay (PCA) in order to find the interaction and correlation between the WT and the deletion backgrounds. While conducting this the article states that the deletion of a paralog in a pair affects the other duplicate directly. Based upon this statement the article argues that one duplicate has a function in the PPI network that cannot be compensated by or “that is not dependent on the other copy”. While conducting and examining transcript abundance in yeast deletion strains the researchers, “did not find any compensating pairs with significant change in mRNA expression in the paralog deletion background.” The reading also states that the PPI of one duplicate would predominate the other due to either higher abundance or affinity and a compensation would result from a deletion of a paralog.
Next the physical interaction between paralogs was measured and it was noticed that dependent pairs are enriched for paralogous heteromers (paralogs with different polypeptides). The dependent duplicate has lower protein abundance than its cognate copy which further decreases when deletion occurs. The article also found data to support the argument that deletion of independent duplicate pairs strikes the fitness more than that of the dependent ones. When analyzing a larger set of paralogs in budding yeast, the researchers have found that deletion of duplicates reduces the fitness more in general. They argue that SSD(small scale duplicates) have a more divergent function than ohnologs and that the deletion of heteromeric paralogs has a larger cost than nonheteromeric paralogs. The same trend has been found in humans which suggests that this in not only limited to yeast. These heteromeric paralogs could therefore be more similar in amino acid sequences and share molecular functions.
Ohnologs in budding yeast were compared to orthologs from Schizosaccharomyces pombe to get more information about their duplication and gain more PPI data. Ohnologs forming paralogous heteromers are more likely having a homomeric ortholog, suggesting that dependency of paralogs might be caused by a homomeric status of the ancestral protein. The article suggests that the duplication of homomers is a mechanism for the evolution of paralogous heteromers. The results show that paralogous genes compensate each other’s loss in PPI networks which adds to robustness of cellular functions. However, many paralogs have an opposite effect which increase the mutational fragility when deletion of a paralog occurs. The results partially explain why duplication of homomers increase the complexity of PPI and often making it more susceptible to mutations. Paralogous genes can compensate each other’s loss in PPI network. However, when deletion of the paralog occurs, it increases the mutational fragility. Duplication of the paralog increases the complexity of protein interactions and also increases susceptibility to mutations.
Opinion:
The article uses good reasoning to convey its
point of view that gene duplication imparts fragility and not
robustness in the yeast PPI network. The data throughout this article
uses good visual evidence and distribution of colors to prove this
specific point. The figures in this paper give a clear and structured
overview of the main gist. The paper did a good job summarizing the
huge amount of information into organized sections ranging from
functional fates of paralogous genes to their reactions among each
other. The figures provided secure the reader with enough data to
represent their point, however some contradictions have not been
explained such as the duplication in homomers which increases the
complexity of PPI and increases the possibility of mutations. Further
investigation has to be pursued in in this field.
Figure Summary:
Fig. 1:
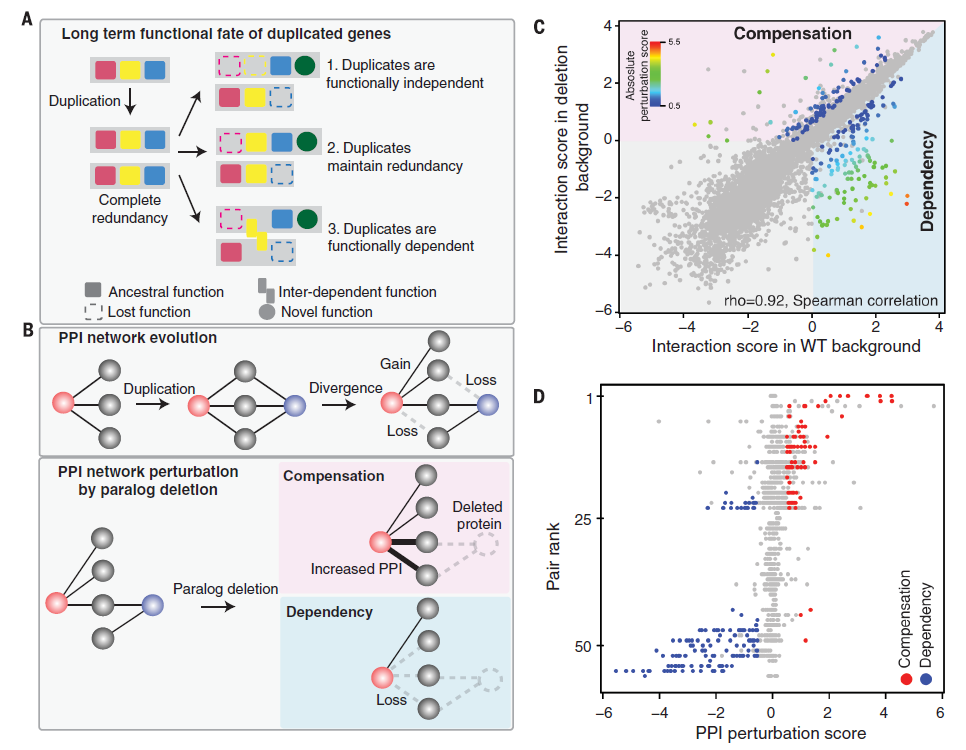
Figure 1 shows the long term functional fate of
duplicated genes and their connection to the PPI network evolution.
Over time the duplicated genes can either be functionally independent,
maintain redundancy or be functionally dependent. However, the
duplicates that maintain redundancy seem to have a functional overlap
which allows a paralog to compensate in case of loss of function.
112 proteins were mapped in the wild type and deletion
background of their cognate copy. The duplication of a gene can
diverge and either lead to a gain or loss in the PPI network
evolution, whereas a paralog deletion either leads to an increase in
PPI (compensation) or losses in PPI (dependency). The interaction
score between wild type and deletion backgrounds was plotted. The
Interaction score in WT background is on the x axis and Interaction
score in the deletion background is on the y-axis. The spearman value
of 0.92 suggests that there is almost a linear correlation between
these two. The grey points show data points that are not statistically
significant. The dependency region (also more wide spread) seems to
have a higher absolute perturbation average than the compensation
region, suggesting that dependent genes have a higher probability of
encountering a mutational change. The data in the next graph(1D)
supports this argument, which clearly shows that compensation has a
higher PPI perturbation score.
Fig 2:
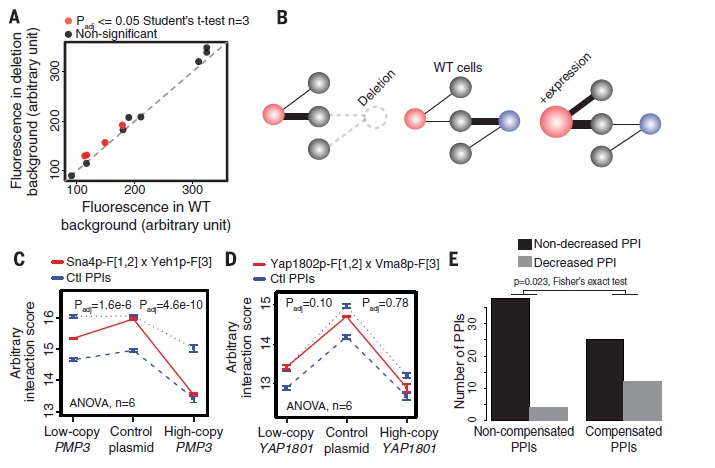
In figure 2A the protein abundance was measured. Three significant changes were marked. This figure argues that if the protein abundance in the WT background increases so does the protein abundance in deletion background (almost linear correlation) The compensation may result from mutual exclusion in WT cells. Deletion of one paralog results in functional compensation(no change in expression), whereas the overexpression of the compensating paralog results in a decreasing interaction involving the other paralog. The interaction of proteins is decreased upon overexpression of PMP3 and YAP1801. As seen the number of PPIs in non-compensated PPIs is way higher for non-decreased PPIs than decreased PPI’s. The same goes for compensated PPI’s(low p-value shows significant difference). The compensated PPI’s tend to be affected more by the overexpression of the compensating paralog, resulting in a decreased PPI number.
Fig 3:
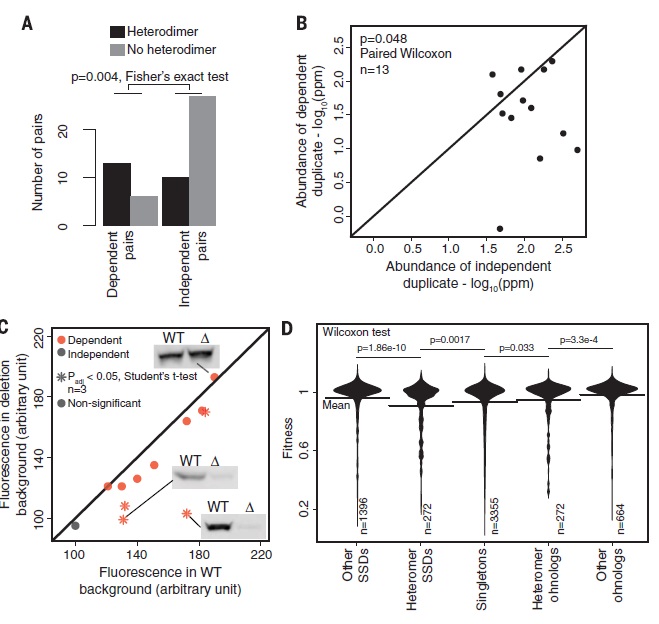
Fig. 3 examines the physical interactions between paralogs which might be a key factor in stabilizing or regulating partners. Dependent pairs have an almost 2 fold higher number of pairs in heterodimer (paralogs that physically interact and are made of two polypeptides) compared to no heterodimer. In independent pairs the pair number of paralogs with no heterodimers are substantially higher than that of the paralogs with a heterodimer(low p-value show significant difference). There is no linear correlation between the abundance of the dependent and independent duplicate. The scattered graph shows a higher protein abundance in independent duplicates than in dependent duplicates. The p value is low enough to indicate that there is a significant difference. The fluorescence in the WT background shows an increase in protein abundance in dependent pairs and a decrease in independent pairs with rising fluorescence intensity in the background. The fitness depends on what gene has been deleted. The deletion of heteromer SSD, Singletons and that of heteromer ohnologs reduces the fitness the most (all measurements showing significant difference).
Fig 4:
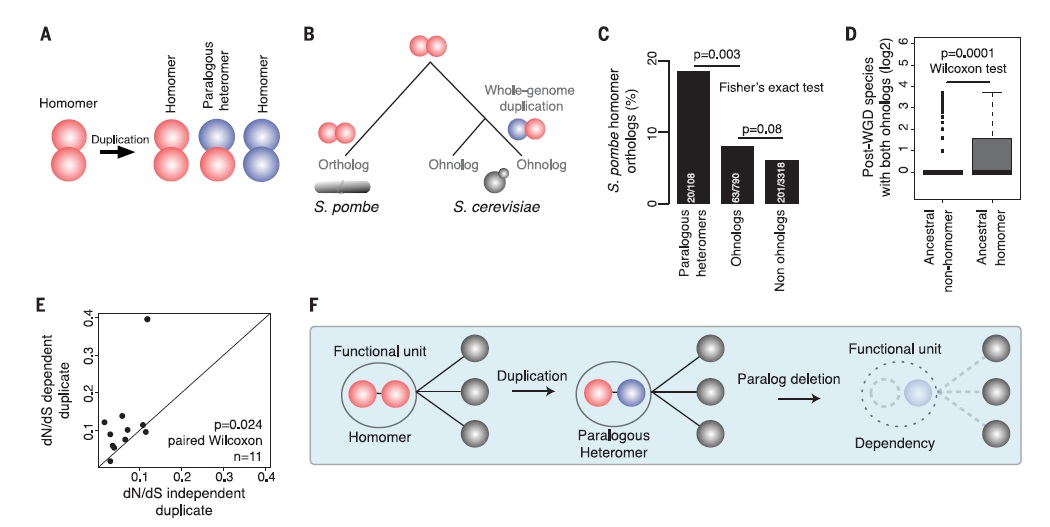
Fig. 4 describes the possibility of duplicates evolution into a heteromer or homomer after duplication. A heteromer might have evolved from the duplication of the same ancestral homomer. Ohnologs from S.cerevisae have been compared to orthologs in S.pombe. The percentage of paralogous heteromers form S.pombe homomer orthologs is about twice as high when compared to ohnologs and non ohnologs(para. Heterom. Approx. 20%). The p-values indicate that there is significant difference between the measurements. This graphical representation suggests that paralogous heteromers are more likely to have diverged from orthologs that come from ancestral homomers. The number of post-WGD species with both ohnologs is significantly higher in the ancestral homomer than in the ancestral non-homomer. The dependent duplicates evolve faster than the independent duplicates, which can be seen by the non-linear correlation and the scattering. The homomers can lead to a functional unit of a paralogous heteromer that is dependent when duplicated. A paralog deletion affects the functional unit and thus leading to a breakdown of the PPI.
Refrences:
Diss, G. (feb 2017). Gene duplication can impart, fragility not robustness, in the yeast protein interaction network.
Email Questions or Comments: pharndt@davidson.edu