Immunological Investigation ofChlamydomonas Malic Enzyme
Amy Lindstrom
Honors Thesis Report
Spring 1997
ABSTRACT
Adhesion proteins (AP) found on the surface
of flagella have been shown to aid in the cytoadherence mechanism of the
single celled parasite Trichomonas vaginalis. One AP, AP65 (Alderete,
et al. Molecular Microbiology 17: 69-83, (1994), resembles
malic enzyme (ME) and anti-AP65 antibodies recognize ME found in T. vaginalis
and chicken liver. ME is an evolutionarily well-conserved protein found
in a variety of organisms. However the presence of this metabolic enzyme
on the surface of T. vaginalis flagella is unusual. To see if ME
is a common feature found on the flagella of other organisms we decided
to probe for this enzyme in Chlamydomonas reinhardii, the model organism
for flagellar research. Cell lysates of C. reinhardii and chicken
liver (purchased from Sigma) were separated on SDS-PAGE for the purpose
of performing an immunoblot analysis. Chicken ME was detected, but C.
reinhardii ME was not. These results suggested that the epitope(s) for
the anti-AP65 antibodies was not present in C. reinhardii ME. A search
of Genbank for ME amino acid sequences from different species was performed
to detect evolutionarily conserved domains. The multiple amino acid sequences
were aligned in order to predict the location of the epitope of the anti-AP65
antibodies.
INTRODUCTION
The single celled eukaryote Chlamydomonas reinhardii is a model organism
for studying the biology and genetic features of photosynthetic pathways
and flagellar structure. C. reinhardii has two flagella located at
the anterior end of the cell; enclosed by a cell wall are the nucleus, visible
nucleolus, chloroplasts, mitochondria and other essential structures (Figure
1).
Figure 1: Schematic representation of the green alga, C. reinhardii. The major morphological features have been identified (adopted from Johson et al., 1993).
Chlamydomonas reinhardii Flagella
C. reinhardii has
served as a molecular model for flagellar and ciliary movement of eukaryotic
cells. For example, C. reinhardii and sperm flagella, along with
cilia that line the respiratory passages, share the typical (9 + 2) microtubule
structure found in most flagellated or ciliated organisms (Figure 2).
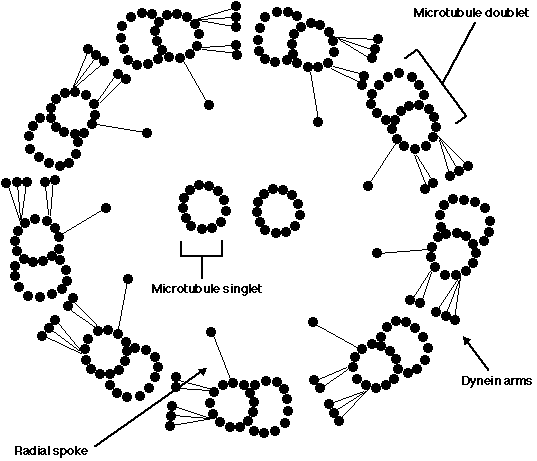
Figure 2: A cross sectional diagram of the axoneme, the (9+2) microtubule structure (adopted from Blair et al., 1992). The nine doublets form a ring around the two microtubule singlets. The radial spokes and dynein arms contribute to maintaining this structure and influencing its movement.
As shown in the diagram, nine microtubule doublets form a ring around two single microtubules. Each of the nine microtubule doublets is composed of thirteen protofilaments arranged in a circle which is adjacent to another ten protofilaments that form a half circle next to it. A microtubule singlet consists of thirteen protofilaments arranged in a circle. Each protofilament is comprised of alpha tubulin/beta tubulin heterodimers which are stacked on top of each other. To help maintain the (9+2) arrangement of microtubules, a radial spoke protrudes from each doublet connecting the outer circle to the central microtubules. In addition, five dynein side-arms extend from each doublet and influence the conjugative movement of this (9 + 2) microtubule structure.
Over 200 proteins contribute to the flagellar structure and function of C. reinhardii (Lefebvre et al., 1980). Of these 200 proteins, approximately 40 have been characterized, all of which are involved in the structure and function of the axoneme, the (9+2) microtubule structure. A membrane encloses the axoneme giving the flagella its cylindrical shape. A large number of flagellar proteins remain uncharacterized, however, some of which are closely associated with the membrane. One major group of these uncharacterized proteins are the extracellular glycoproteins, called agglutinins, present on the flagellar tips during gamete agglutination.
C. reinhardii
has two morphologically indistinguishable mating types called plus and minus.
Each mating type has its own set of distinct extracellular glycoproteins
that aid in gamete recognition (Harris, 1989). Gametes cannot mate with
the same mating type and can only form a diploid zygote by fusing with a
cell of the opposite mating type. These flagellar glycoproteins facilitate
the actual fusion process by binding to each other, thus bringing the cell
bodies into close contact and placing the gametes in an appropriate position
suitable for cell fusion. Therefore, extracellular flagellar proteins serve
an important role in the mating process. Hence other unidentified flagellar
proteins probably exist, which could contribute to equally fundamental processes.
Flagellar Proteins
From Other Organisms
An unexpected discovery recently reported the presence of metabolic enzymes
on the extracellular surfaces of microbes and their flagella (Camara et
al., 1994; Joe et al., 1994). These enzymes have been shown to
have many functions other than metabolism. Usually the location of a protein
will determine its function. Therefore, when these normally intracellular
metabolic enzymes are moved onto the extracellular surface of the cell,
their functional role changes. Instead of aiding in cellular metabolism,
these extracellular proteins serve as receptors or adherence molecules (Lottenberg
et al., 1992; Pancholi and Fischetti, 1992; 1993; Vacca-Smith et
al., 1994). A recent study by Alderete et al. (1995) revealed
the presence of malic enzyme (ME) on the flagella of the parasite Trichomonas
vaginalis.
The single-celled, amitochrondriate eukaryote T. vaginalis is a medically important parasite which adheres to the vaginal epithelial cells (VEC) of women. Its morphology includes four anteriorly positioned flagella, a fifth flagellum facing posteriorly surrounded by an undulating membrane and an axostyle extending past the base of the cell (Figure 3).
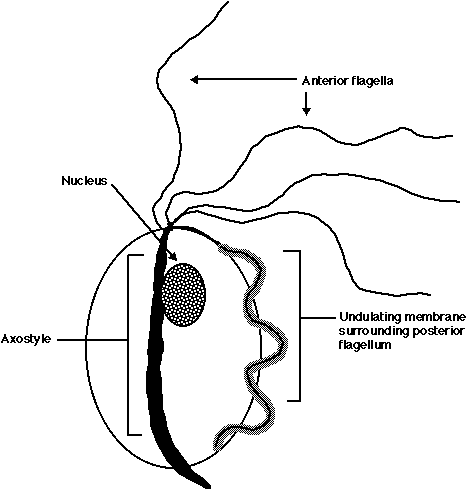
Figure 3: Schematic representation of the parasite T. vaginalis. The major morphological features have been identified (adopted from Ash LR and Orihel TC, 1980).
Alderete et al. have characterized four adhesion proteins (AP) found on the surface of T. vaginalis's flagella. These APs enable T. vaginalis to bind to the VECs of its host. On one of the APs, Alderete et al. identified a receptor site that binds to a ligand on the VECs. This observation suggested that cytoadherence by T. vaginalis occurs in a ligand/receptor fashion; the APs serve as receptors that bind to a specific site (ligand) found on the surface of VECs (Figure 4).
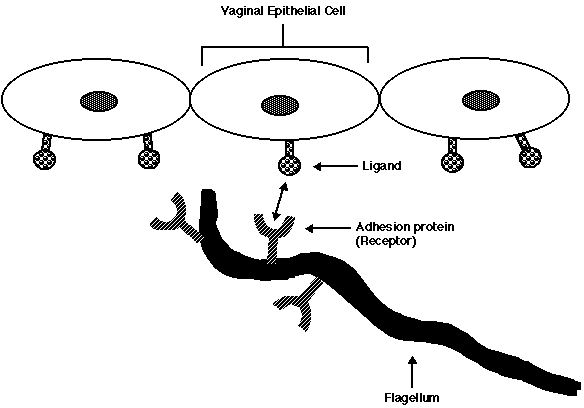
Figure 4: Diagramatic interpretation demonstrating how the adhesion proteins on the flagella of T. vaginalis serve as receptors. The receptors attach to a ligand present on the vaginal epithelial cell and enable the parasite to adhere to its host.
The parasite's ability to maintain cytoadherence
in an environment being constantly flushed with vaginal secretions "is
a critical step in infection" (Alderete et al., 1995). By characterizing
the APs, Alderete et al. could grasp a better understanding
of T. vaginalis's ability to maintain cytoadherence. Therefore, they
isolated a cDNA encoding one of the APs, AP65. Alderete et al. sequenced
the cDNA encoding AP65 and then searched for homology among other proteins.
Surprisingly, they discovered that AP65 has a high degree of homology with
malic enzyme. In fact, Alderete et al. found a "38 % amino acid
identity and 54 % nucleotide identity of AP65 with ME"(Alderete et
al., 1995).
Malic Enzyme
There are two classes of enzymes which are commonly referred to as ME. One
of the enzymes is the NAD+-dependent malate dehydrogenase that is located
in the mitochondria. It plays an important role in the citric acid cycle
by oxidizing malate to form oxaloacetate and reducing NAD+ to NADH (Figure
5).
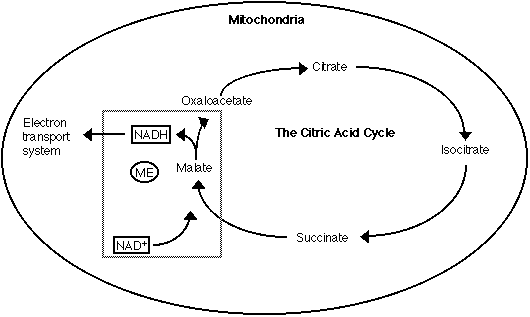
Figure 5: A brief overview of the Citric Acid Cycle
The boxed area represents NAD+- dependent dehydrogenase's role of oxidizing
malate to form oxaloacetate and NADH. The NADH enters the electron transport
system where it aids in the production of ATP (adopted from Stryer, 1995).
The NADH enters the electron transport system where it aids in the production of ATP. The other enzyme, NADP+-dependent malate dehydrogenase, is a scavenger enzyme found in the cytosol. The primary function of this class of ME is the decarboxylation of malate to form pyruvate, NADPH and CO2 (Figure 6).
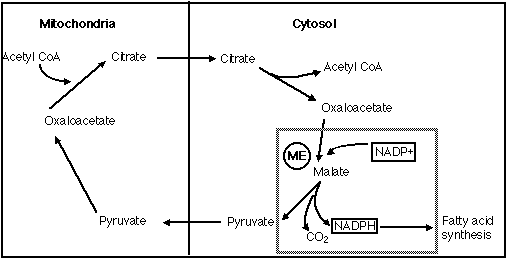
Figure 6: The boxed area demonstrates NADP+-dependent dehydrogenase's role of decarboxylating malate to form pyruvate, CO2, and NADPH. The NADPH aids in the production of fatty acids (adopted from Stryer, 1995).
NADPH then serves as a reducing agent necessary for fatty acid synthesis. Fatty acids not only are an excellent energy source, but also serve as structural units in the phospholipid bilayer. Thus this NADP+-dependent dehydrogenase is essential for maintaining the metabolic and structural integrity of a cell.
Both the NAD+ and NADP+-dependent malate dehydrogenases are found in bacteria, plants and animals. Because of ME's ubiquitous nature, there was reason to suspect that it could be found in C. reinhardii. In fact, cell lysates from C. reinhardii were run on a column to obtain a relatively purified sample of ME. This sample was analyzed by polyacrylamide gel electrophoresis and stained for activity. The gel revealed that ME is present in this green alga (Williamson et al., personal communication).
Not only is ME widespread, but it is a relatively
well conserved protein found in a variety of organisms. It would be reasonable
to expect that antibodies specific to ME from one organism would bind to
ME from another. In fact, both the polyclonal and monoclonal anti-AP65 antibodies
cross reacted with ME from T. vaginalis and chicken liver, suggesting
that the epitope sites are highly conserved between these two organisms.
Alderete et al. had demonstrated that ME, normally classified as
a metabolic enzyme, is found on the flagella of T. vaginalis. Therefore,
ME's presence on the surface of trichomonal flagella and the previously
discussed knowledge of metabolic enzymes found on other microbial surfaces
lead to an interesting question: Is malic enzyme a characteristic of C.
reinhardii's flagella, the model organism used for flagellar research?
Immunoblots were designed to determine if the anti-AP65 monoclonal and polyclonal
antibodies reacted with ME found in protein extracts from C. reinhardii.
Assuming positive results, immunofluorescence would be done on intact C.
reinhardii cells. This technique would enable us to visualize the location
of ME on C. reinhardii. We could then determine if ME is part of
the flagellar structure or if it is only found inside the cell body. The
results and the significance of the data are discussed in this paper.
MATERIALS AND METHODS
Preparation of C. reinhardii cells
Wild-type cells (cc125, from culture collection at Duke University) were
grown in Erlenmeyer flasks in liquid media (Tris-acetate-phosphate, TAP;
Harris, 1989), under constant illumination at ambient temperature. Cells
were recovered from culture medium by centrifugation at 3,800 g for five
minutes, resuspended in TEM buffer (0.1 M Tris, 5 mM EDTA, 1 mM 2-mercaptoethanol,
pH 7.6), and stored at -70 oC until needed.
Cellular fractionation of tissue
Cells were removed from the -70 oC freezer
and allowed to thaw at room temperature. They were centrifuged at 16,500
g for ten minutes. Ammonium sulfate was added to the supernatant to 40 %
saturation. The sample was stirred for twenty minutes at 4 oC and centrifuged
at 16,500 g for ten minutes. Ammonium sulfate was added to the supernatant
to 65 % saturation and then centrifuged again at 16,500 g for ten minutes.
Afterwards, the supernatant was discarded and the pellet was resuspended
in 4 ml TEM buffer.
Assays for enzyme activity
ME activity was monitored at 340 nM using
a Beckman Model 35 spectrophotometer following the procedure of Geer, et
al.(1980). Twenty microliters of the enzyme prep was added to 1 ml of
malic enzyme assay buffer (0.133 M Tris, 0.34 mM NADP, 1 mM MnCl2, pH 7.9)
in a 1 ml cuvette and incubated in a 30 oC water bath for 2 minutes. The
reaction was initiated by adding 10 ul of 10 mM malate.
Affinity chromatography
The resuspended pellet was desalted over a Sepharose G-50 column. Fractions
containing ME activity were combined and pumped over a Matrix Blue A column.
This column is composed of Matrix Blue F3G-A covalently attached to cross-linked
Sepharose by the triazine coupling method (Amicon). The Matrix Blue F3G-A
dye has been previously shown to bind to malic enzyme which could be eluted
with 1.5 M KCl (Amicon). Aliquots (2.9 ml) were collected and assayed for
ME activity. Tubes containing ME activity were combined and concentrated.
The final activity in 20 ul was 0.081 units (1 unit = 1 uM NADP reduced
per minute). An activity of 0.072 units of C. reinhardii was used
in the western blots (Figures 8-11). An activity of 0.101 units/5 ul of
chicken ME was used in the western blots (Figures 7-11).
Procedure for western blot analysis
A pre-cast 10 % SDS-polyacrylamide gel was assembled in a Mini-PROTEAN II
cell that contained 300 milliliters of electrode buffer (Biorad ). The protein
samples were loaded onto the gel and electrophoresed at 200 V for 45 minutes.
Afterwards, the gel was equilibrated in transfer buffer (25 mM Tris base,
1.9 M Glycine, 70 % water and 20% methanol) for 15 minutes. The transfer
unit then was assembled, placed into the transfer chamber and run at 100
V for 1 hour. The nitrocellulose membrane was then blocked in Blotto (7%
dry milk, 1 X Tris-Saline (9.98 mM Tris base, 1.5 M NaCl), 0.1 % Tween 20
and 0.1 % antifoam A) for one hour at room temperature with gentle shaking.
The Blotto was poured out and the membrane was rinsed with Tris-Saline.
The membrane was incubated overnight at 4 oC in 5 milliliters of Blotto
that contained either a 1: 10 dilution of the monoclonal antibody or a 1:
20 of the polyclonal antibody. The membrane was washed two times for five
minutes each with Tris-Saline. The membrane probed with the mouse monoclonal
antibody was incubated in 10 milliliters of Blotto that contained a 1:2,500
dilution of horseradish peroxidase-conjugated goat anti-mouse antibody for
one hour at room temperature. The membrane incubated with the polyclonal
antibodies contained a 1: 5,000 dilution of horse radish peroxidase-conjugated
goat anti-rabbit antibody for one hour at room temperature. Both membranes
were washed three more times, five minutes each with Tris-Saline. The horse
radish peroxidase substrate, True Blue (Kirkegard and Perry Laboratories),
was added to detect the antibody binding site on the gel. The gels were
photographed on Foto/Analyst Archiver (Fotodyne).
RESULTS
Probing for chicken malic enzyme
The purpose of Figure 7 was to verify that ME from chicken liver reacted
with the monoclonal anti-AP65 antibodies that bind to ME from T. vaginalis.
Monoclonal antibodies are specific to one epitope; polyclonal antibodies
are a group of immunoglobins all specific to a particular antigen, yet the
different antibodies bind to separate epitopes located on this antigen.
Therefore if the monoclonal anti-AP65 antibodies reacted with chicken ME,
we felt it would be safe to assume that the polyclonal anti-AP65 antibodies
would too. A single band between the 80 kDa and 50.9 kDa MW markers appeared
in lanes 2, 4, 6, and 8 (Figure 7).
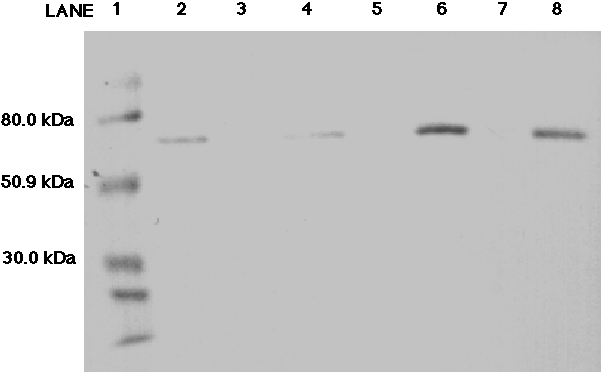
Figure 7. Western blot analysis probing for malic enzyme from chicken liver using the anti-AP65 monoclonal antibody. Five, ten, thirteen and thirteen (unboiled sample) microliters (activity of 0.101 units/5 ul) of chicken malic enzyme were loaded into lanes 2, 4, 6 and 8 respectively. The molecular weight ladder was loaded in lane 1. The 80.0 kDa, 50.9 kDa and 30.0 kDa markers have been labeled. A clear band appears in lanes 2, 4, 6 and 8 between the 80.0 kDa and 50.9 kDa markers.
We found that an activity of 0.101 units of chicken
ME (lane 2) was enough to give a clear, distinct band that could serve as
a positive control.
ME is a homo-tetrameric enzyme, each of the four
subunits between 60-70 kDa in size depending on the species. The band which
appears in this blot has migrated just below the 80 kDa MW marker, indicating
that it is in the expected 60-70 kDa range. In addition, an unboiled sample
(lane 8) was tested to see if the samples must be boiled first in order
for the proteins to resolve properly in the gel. Lane 8 demonstrates that
boiling is not necessary for disassociation of the ME subunits.
Probing for C. reinhardii malic enzyme
The subsequent two western blots probed
for both chicken ME and C. reinhardii ME with either the monoclonal
antibody (Figure 8) or the polyclonal anti-AP65 antiserum (Figure 9).
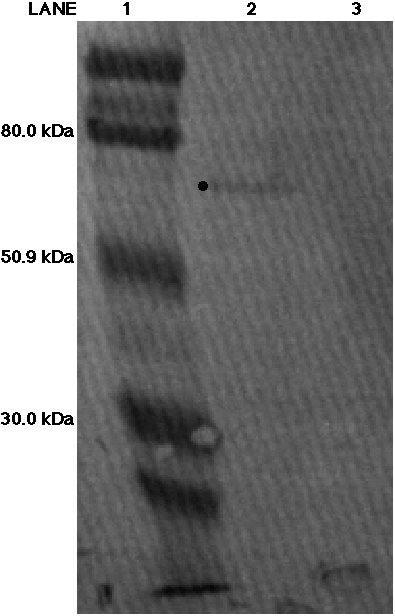
Figure 8. Western blot analysis probing for malic enzyme fromChlamydomonas reinhardii using the anti-AP65 monoclonal antibody. An activity of 0.101 units of chicken ME was loaded into lane 2 and an activity of 0.072 units of C. reinhardii ME was loaded into lane 3. The molecular weight ladder was loaded into lane 1. The 80.0 kDa, 50.9 kDa and 30.0 kDa markers have been labeled.A band appears in lane 2 (identified by dot) but no band is present in lane 3.
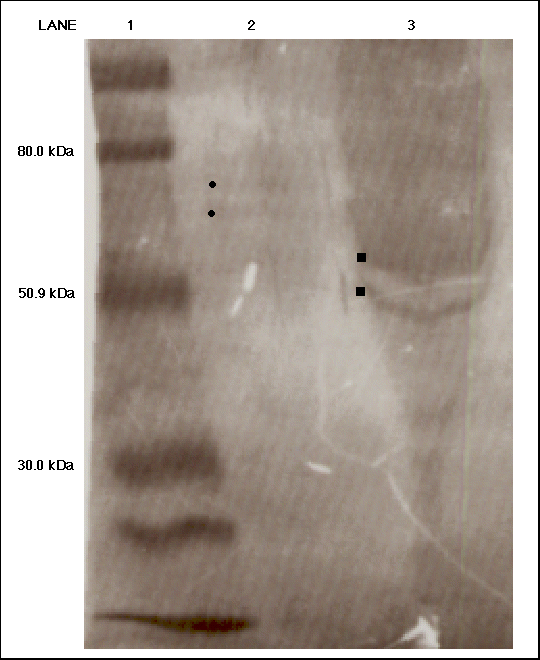
Figure 9. Western blot analysis probing for malic enzyme from Chlamydomonas reinhardii using the anti-AP65 polyclonal antibody. An activity of 0.101 units of chicken ME was loaded into lane 2 and an activity of 0.072 units was loaded into lane 3. The molecular weight ladder was loaded into lane 1. The 80.0 kDa, 50.9 kDa and 30.0 kDa markers have been labeled. Two faint bands (identified by dots) appear in lane 2 and two potential distorted bands appear in lane 3 (identified by squares).
In Figure 8, a faint band appeared just below the 80 kDa MW marker in the chicken lane, but the C. reinhardii lane remained blank. The MW ladder in both Figures 8 and 9 are distorted suggesting that the potassium chloride salt in the C. reinhardii samples is preventing a linear migration of the bands. The potassium chloride was used to elute ME from the column during affinity chromatography and the sample was never dialyzed to remove the salt. In Figure 9, two faint bands appear in lane 2 loaded with chicken ME. They have migrated close to ME's molecular weight range, suggesting that perhaps two forms of ME are present in the chicken sample. Similarly, what appears as two bands are present in the C. reinhardii lane of Figure 9. However, the salt has especially disturbed the proper migration in this lane, thus preventing us from making any definite conclusions about these two bands. In both Figures 8 and 9, the migration of the bands are distorted, presumably due to the presence of potassium chloride found in the C. reinhardii samples. During electrophoresis, the salt spread across the gel disrupting the proper migration of the protein bands (Deutcher, 1990). To obtain better results, we performed another western blot using a dialyzed C. reinhardii sample. Dialyzing would get rid of the excess salts and permit a better migration of the bands. Also, by removing the potassium chloride, we would see if the antibodies could better detect the chicken ME. In addition, the band which appeared in lane 2 of Figure 7 was fainter than the band present in the chicken lane of Figure 8; both lanes had an activity of 0.101 units of chicken ME loaded on the gel. The faintness of the band in the chicken lane of Figure 8 suggested that the antibody signal as a whole had weakened because the western blot in Figure 8 was incubated with the same monoclonal antibody solution as the one in Figure 7. Therefore the western blot in Figure 7 could have depleted the solution of antibodies, thus causing a weaker signal in the western blot in Figure 8. For Figure 10, an additional 50 microliters from the original monoclonal antibody sample was added to the previously used incubation solution. By dialyzing the C. reinhardii protein sample and increasing the antibody concentration, we hoped to obtain better results in the next two western blots.
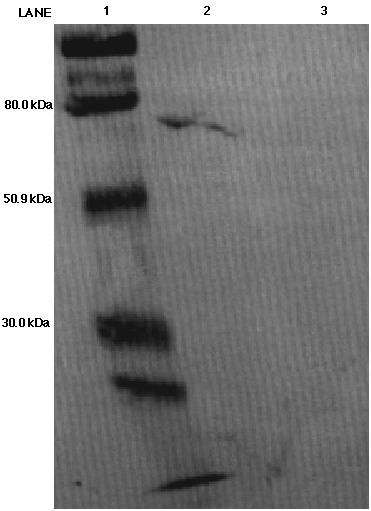
Figure 10. Western blot analysis probing for malic
enzyme from C. reinhardii using the anti-AP65 monoclonal antibody.
An activity of 0.101 units of chicken ME was loaded in lane 2 and an
activity of 0.072 units of C. reinhardii ME was loaded in lane 3.
The molecular weight ladder was loaded in lane 1. The 80.0 kDa, 50.9 kDa
and 30.0 kDa markers have been labeled. A distorted band appears in lane
2, but no band appears in lane 3.
Probing for C. reinhardii malic enzyme
from dialyzed samples
In Figure 10 the western blot was incubated with the anti-AP65 monoclonal
antibody. In the chicken lane, a band appears just beneath the 80 kDa marker
but no band appears in the C. reinhardii lane. The western blot shown
in Figure 11 was incubated with the anti-AP65 polyclonal antibodies. The
MW ladder migrated much better in Figure 11 than in Figures 8, 9 and 10.
However lanes 2, 3 and 5 of Figure 11 suggest that what little salt is present
may still be distorting the migration of the bands. In Figure 11, both the
chicken lane and C. reinhardii lane (lane 3) have no distinguishable
bands, only what appears to be slightly darker smudges running down each
lane. In the unboiled C. reinhardii lane (lane 5) there appears to
be two bands, the smaller one being even with the 50.9 kDa marker and the
larger one located directly above it. However the bands are distorted, making
it difficult to interpret.

Figure 11: Western blot analysis probing for malic
enzyme from Chlamydomonas reinhardii using the anti-AP65 polyclonal
antibody. An activity of 0.101 units of chicken ME was loaded into lane
2 and an activity of 0.072 units of C. reinhardii ME was loaded into lanes
3 (boiled sample) and 5 (unboiled sample). The molecular weight ladder was
loaded into lane 1. The 80.0 kDa and 50.9 kDa markers have been labeled.
DISCUSSION
Due to the small amount of the anti-AP65
antibodies given to us, only a minimum number of western blots could be
performed. After doing the western blot in Figure 10, we ran out of the
monoclonal anti-AP65 antibody, but still had enough of the polyclonal antibodies
for one more blot. Based on the results obtained in Figures 8-11, neither
the anti-AP65 monoclonal nor the polyclonal antibodies bound to ME from
C. reinhardii. These negative results could suggest that the epitope
site(s) to which the monoclonal and polyclonal antibodies are specific may
not be present on C. reinhardii ME. Perhaps the amino acid sequence
of ME from this green alga varies greatly from those in most other organisms
and the epitope site would not be found on C. reinhardii ME. Whatever
the reason, the anti-AP65 antibodies failed to bind to ME from C. reinhardii.
Before attempting one last western blot with the anti-AP65 polyclonal antibodies, I decided to do a search of Genbank looking for sequences of NADP-dependent malate dehydrogenases from different species. Using MacDNAsis, I constructed a multiple amino acid sequence alignment of ME from a variety of organisms. I then studied the alignment, searching for conserved regions found throughout MEs from the array of organisms. I paid particular attention to conserved regions which contained at least 8 amino acids aligned consecutively, for the average epitope site is between 8-12 amino acids in length (Churchill et al., 1994; Lind et al., 1992; Watson et al., 1992; Wilson et al., 1992). A few conserved regions were found, however most were between 4-6 amino acids long. Only one conserved region was found, among all the organism except for one (Flaveria trinervia), that could potentially serve as an epitope site. This site is located between amino acids 267-275, making it 9 amino acids long (Figure 12).
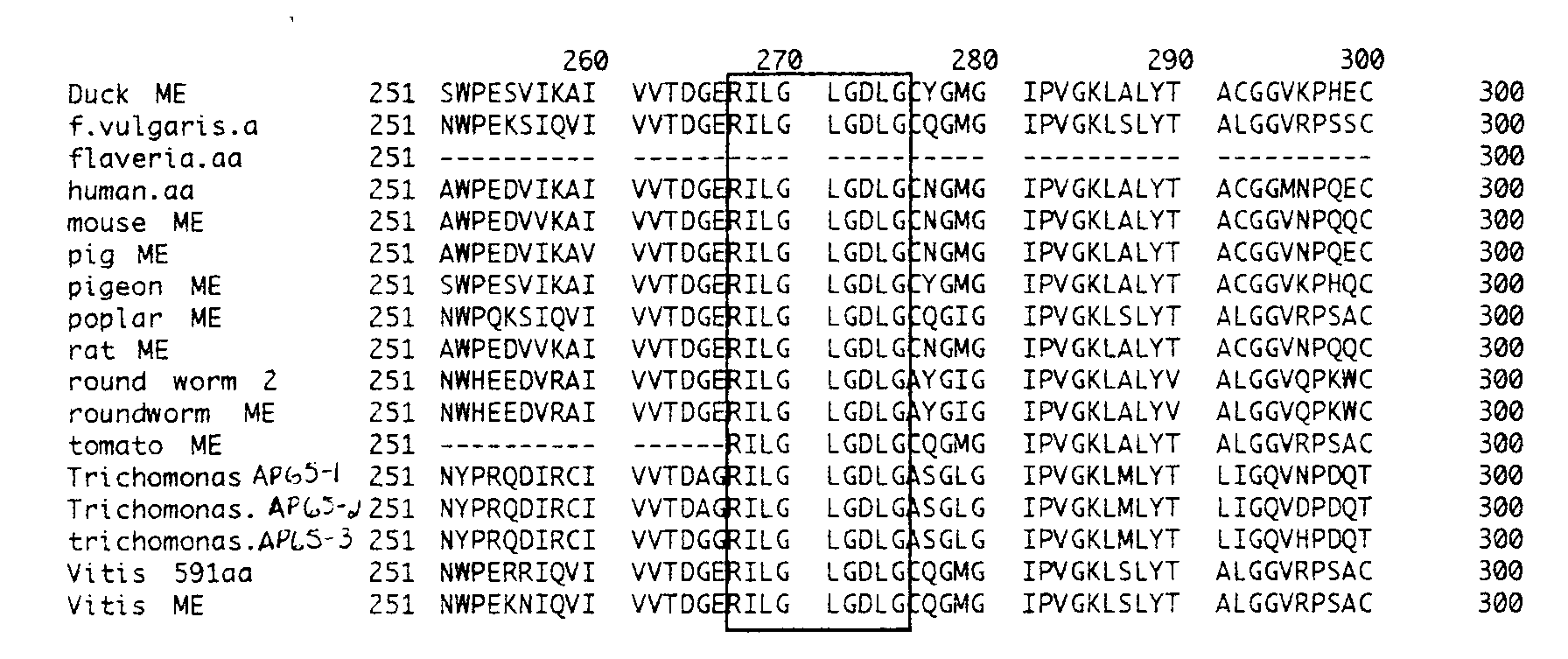
Figure 12: The boxed area indicates the site of the possible epitope for the anti-AP65 antibody. The site is between amino acids 267- 275, making it nine amino acids long.
The length of this conserved region makes it a strong candidate for the epitope site of the AP-65 monoclonal antibodies. Malic enzyme from C. reinhardii has not been sequenced. Therefore there is no way of knowing whether this potential epitope site is conserved on ME from this green alga. However based on the results in the western blots, this nine amino acid long sequence appears not to be present on malic enzyme from C. reinhardii.
REFERENCES
Alderete JF, O'Brien JL, Arroyo R, et al. Cloning and molecular characterization
of two genes encoding adhesion proteins involved in Trichomonas vaginalis
cytoadherence. Mol Microbiol 1995; 17: 69-83.
Ash LR, Orihel TC. Atlas of Human Parasitology. Chicago: American Society
of Clinical Pathologists Press, 1980.
Blair DF, Dutcher SK. Flagella in prokaryotes and lower eukaryotes. Gen
and Dev 1992; 2: 756-767.
Camara M, Boulnois GJ, Andrew PW, Mitchell TJ. A neuraminidase from Streptococcus
pneumoniae has the features of a surface protein. Infet Immun 1994;
62: 3688-95.
Churchill ME, Stura E, Pinilla C, et al. Crystal structure of a peptide
complex of anti-influenza peptide antibody Fab 26/9. J Mol Biol 1994; 241:
534-56.
Deutcher MP, ed. Guide to Protein Purification. Methods and Enzymology Series.
San Diego: Academic Press, 1990. Vol. 5: 433.
Geer BW, Krochko D, Oliver MJ, Walker VK, Williamson JH. A comparative study
of the NADP-malic enzymes from Drosophila and chicken liver. Comp
Biochem Physiol 1980; 65: 25-34
Harris EH. The Chlamydomonas sourcebook: a comprehensive guide to
biology and laboratory use. San Diego: Academic Press Inc., 1989.
Joe A, Murray CS, McBride BC. Nucleotide sequence of a Porphyromonas
gingivalis gene encoding a surface-associated glutamate dehydrogenase
and construction of glutamate dehydrogenase-deficient isogenic mutant. Infect
Immun 1994; 62: 1358-68.
Johnson KA, Rosenbaum JL. Flagellar regeneration in Chlamydomonas: a
model system for studying organelle assembly. Trends in Cell Bio 1993; 3:
156-160.
Lefebvre P, Silflow C, Wieban E, Rosenbaum J. Cell 1980; 20: 469-77.
Lind S, Lindberg A. Epitope size, specificity and equilibrium constant for
4 mABs binding to the 0:4 polysaccharide antigen of salmonella serogroup
B bacteria. Mol Immunol 1992; 29: 1013-23.
Lottenburg R, Broder CC, Boyle MDP, et al. Cloning, sequence analysis,
and expression in Escherichia coli of a streptococcal plasmin receptor.
J Bacteriol 1992; 174: 5204-10.
Pancholi V, Fischetti VA. Glyceraldehyde-3-phosphate dehydrogenase on the
surface of group A streptococci is also on ADP-ribosylating enzyme. Proc
Natl Acad Sci USA 1993; 90: 8154-58.
Stryer L. Biochemistry. New York: W. H. Freeman and Company, 1995.
Vacca-Smith AM, Jones CA, Levine MJ, et al. Glucosyltransferase mediates
adhesion of Streptococcus gordini to human endothelial cells in
vitro. Infect Immun 1994; 62: 2187-94.
Watson JD, Witkowski J, Gilman M, Zoller M. Recombinant DNA. New York: W.
H. Freeman and Company, 1992.
Wilson IA, Niman HL, Houghten RA, et al. The structure of an antigenic
determinant in a protein. Cell 1984; 37: 767-78.
![]()
![]()
© Copyright 2000 Department of Biology, Davidson
College, Davidson, NC 28036
Send comments, questions, and suggestions to: macampbell@davidson.edu