This web page was produced as an assignment for an undergraduate course at Davidson College.
HIV Overview and Life Cycle
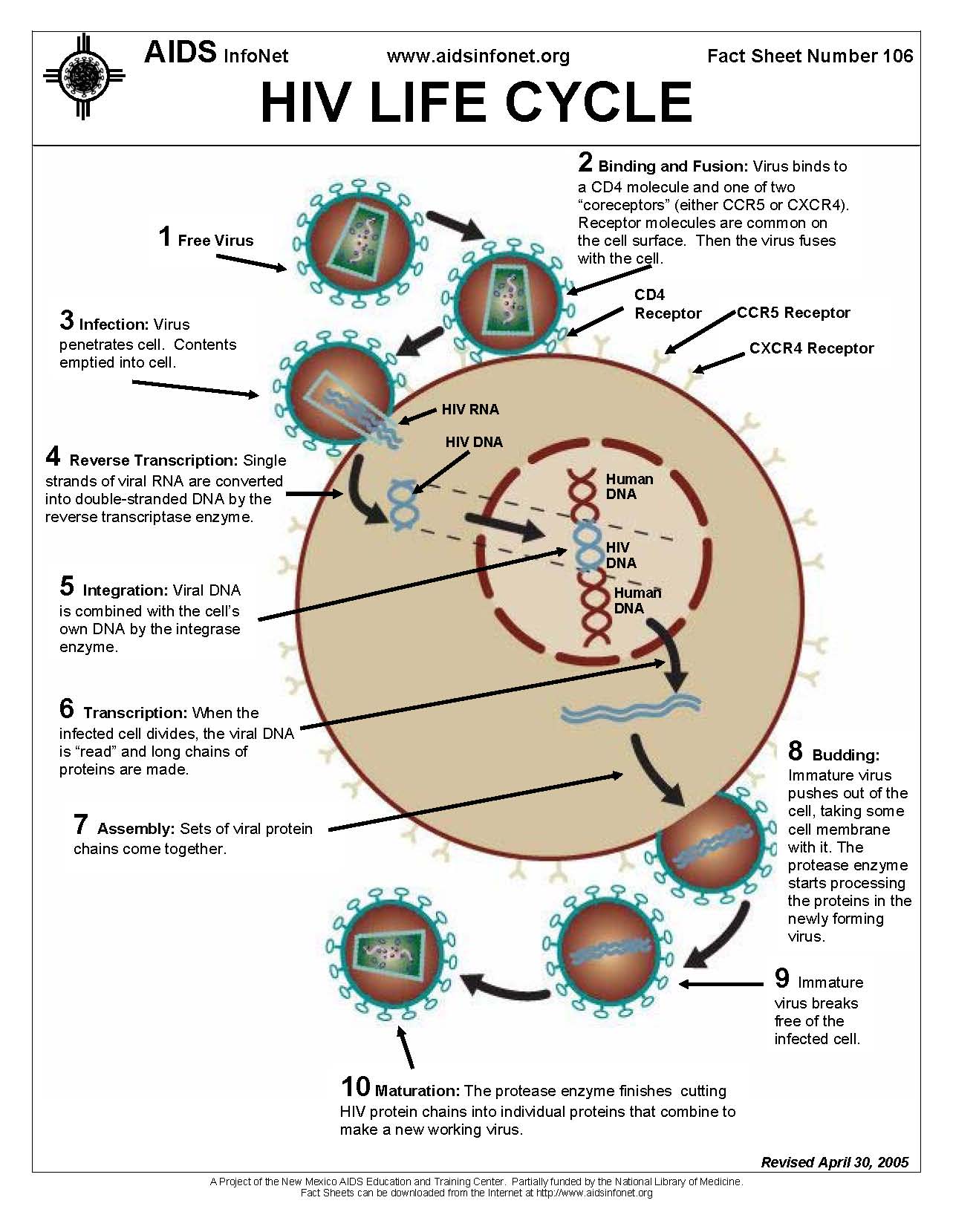
Figure 1. The figure above displays the HIV life cycle in a 10-step process. The figure is from the AIDS InfoNet and the University of New Mexico and can be accessed online here.
The gradual progression of symptoms from HIV earned it a place in the category of similarly slow lentiviruses. Two copies of the RNA genome are stored inside the nucleocapsid along with integrase, protease and reverse transcriptase to facilitate its retroviral activity after invasion of a target cell. The RNA contains nine genes, as well as long terminal repeat sequences (LTR) that help incorporate the genetic instructions of the virus into the cell's DNA. The capsid is contained inside an envelope stolen by the virus particle upon budding from the host cell. One of the HIV genes, env, codes for the protein gp160, which is subsequently cleaved into gp120 and gp41 (Rubbert et al, 2006). The loose complex of gp120 and gp41, assist the virus in entering target cells. Gp120 binds with high affinity to CD4, specifically, to residues in the V2 region that overlap the region where MHC-II molecules bind (Rubbert et al, 2006). While CD4 predominantly found of CD4+ T cells, it is also expressed to a lesser extent on dendritic cells and macrophages (Janeway et al, 2005). But CD4 alone is not enough for HIV to enter a cell, and in 1995, it was discovered that chemokine receptors served as the necessary co-receptor molecule. CCR5 and CXCR4 were found to be the primary chemokine receptors used by HIV. Gp120 binding to CD4 causes conformational changes in gp120 that create a higher affinity for the co-receptor and further changes seem to allow gp41 to insert a hydrophobic region into the membrane of the target cell (Rubbert et al, 2006).
HIV's dependence on chemokine receptors was reinforced by the discovery if individuals seemingly immune to HIV despite frequent high-risk activities. About 1% of caucasians are homozygous for a CCR5 mutation that prevents CCR5-dependent HIV subtypes from fusing with cell membranes, and most initial infections are caused by these so called M-tropic (monocytotropic) subtypes (Rubbert et al, 2006).
As with most other retroviruses, HIV contains three common genes: gag (group-antigen), pol (polymerase) , and env (envelope) (Rubbert, 2006). Gag and Env code for proteins in the nucleocapsid and viral membrane, while pol codes for reverse transcriptase, among other things (Rubbert, 2006). Six other genes - vif, vpu, vpr, tat, rev and nef - differentiate HIV from other retroviruses. Theses six genes are not necessary for HIV replication in vitro, but they are what give HIV its potent characteristics in vivo (Rubbert et al, 2006). Once inside the cell, viral reverse transcriptase transcribes the RNA into cDNA which is then incorporated into to the host genome by the viral integrase, creating what is known as a provirus. Generally, transcription of the provirus initiates replication, but HIV can also establish dormant infections in cells such as memory CD4 T cells (Janeway et al, 2005). Activated CD4 T cells produce transcription factors such as NFkB that bind to LTRs and thus induce transcription of viral RNA by cellular RNA polymerase (Rubbert et al, 2006). In HIV-infected, unstimulated T cells, the cellular protein Murr1 inhibits the degradation of Ik Ba and thus prevents NFkB from entering the nucleus, thereby preventing HIV replication and creating a reservoir of infection (Rubbert et al, 2006).
The rapid replication of HIV coupled with the relatively high mutation rate that occurs during transcription of the virus makes HIV a particularly hard virus to treat. In an infected body, 10^9 to 10^10 new virus particles are created each day (Janeway et al, 2005). Some tactics of treating HIV have focused on creating protease inhibitors to incapacitate the virus' ability to cleave its polyproteins into functional proteins: resistance to some protease inhibitors has been found in just a matter of days in some tests (Janeway et al, 2005). Current treatments rely on cocktails of viral protease inhibitors and nucleoside analogs (AZT for example). While treatment can prolong the asymptomatic period of the virus, CD4 T cell levels eventually drop. CD4 T cells infected with the HIV virus may be killed directly by the virus, are more susceptible to apoptosis and are targeted for destruction by cytotoxic CD8 lymphocytes (Janeway et al, 2005). When CD4 T cell levels fall below a critical level the body loses its ability to mount an effective adaptive immune response to invaders and the patient is said to have AIDS.
References:
Janeway, Charles A, et al. Immunobiology. 6th ed. New York: Garland Science Publishing, 2005.
Rubbert, A., et al . 2006. Pathogenesis of HIV-1 Infection. In: Hoffmann, C., Rockstroh, J., Kamps, B., editors. HIV Medicine 2006. Flying Publisher. <http://www.hivmedicine.com/index.htm>. (5 May 2007).