This web page was produced as an assignment for an undergraduate course at Davidson College.
Evasion of the Immune System
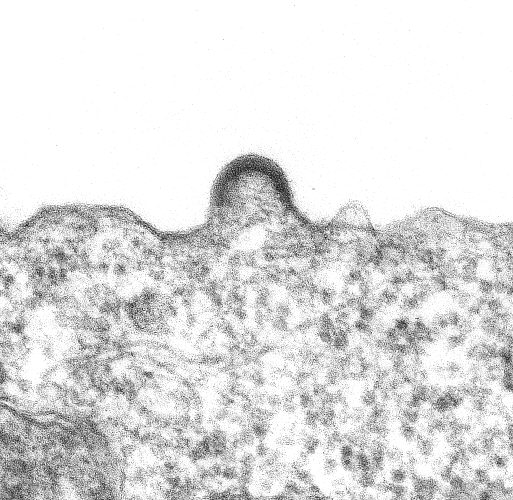
Figure 1. A transmission electron micrograph image of HIV-1 budding from a cultured lymphocyte. The photocan be found at the Public Health Image Library - ID # 947 - CDC/Dr. Edwin P. Ewing, Jr.
HIV uses a wide variety of tactics to evade detection by the immune system at various stages of infection ranging from hiding its presence, to flat out killing the cells that would otherwise seek it out. One if its most effective methods of outsmarting the human immune system comes from a combination of the nature of retroviruses, and the cells that it actively infects. Replicating at a rate of 10^9 - 10^10 new virions each day, with a mutation rate of 3x10^-5 nucleotide per cycle (Janeway et al, 2005) produces several new variations each day within a single infected individual. Not only does this make it incredibley difficult to effectively treat the infection, it makes it just as hard for memory cells, which are specific, to keep up with the virus. Moreover, by infecting memory cells themselves (infected memory cells have a half-life of 44 months), and by infecting macrophages and dendritic cells (which can host the virus without being killed by it), HIV is able to establish reservoirs of infection to continue attacking the host (Janeway et al, 2005).
But HIV has also evolved more direct methods of evasion, especially during the onset of infection when viral titers are lower. One of its more subtle tactics is the opportunistic use of cellular complement inhibitors. When exiting the membrane of a host cell, new virons recruit proteins such as DAF and MCP into their envelopes in order to reduce the chance that they will be killed in plasma before finding a new host cell to infect (Finlay and McFadden, 2006). In a somewhat more high-tech method of cammoflage, the HIV protein Nef plays a role in downregulating the expression of various proteins needed for recognition by potentially dangerous CD8 T cells. Thought to be an adaptor protein, Nef lowers the surface expression of CD4, and several haplotypes of MHC-I by redirecting their transport from the trans-Golgi network (Appay et al, 2000). Interestingly, Nef lowers the expression of MHC-I HLA-A and HLA-B haplotypes, but not of HLA-C or HLA-E haplotypes. This is presumabely because the latter haplotypes are involved in the inhibition of killing by NK cells (Appay et al, 2000). Another gene, Tat, appears to upregulate the expression of Bcl-2 during the early phase of cellular infection, increasing the likelihood that it will receive survival signals (Gougeon and Debatin, 1999).
During later stages of infection, the virus takes a more head-on approach to evasion of the immune system, and turns its efforts on attacking the cells that would kill it. While Tat upregulates Bcl-2 expression initially, establishment of stable Tat-expressing cell lines has been shown to sensitize cells to apoptosis (Gougeon and Debatin, 1999). Similarly, the HIV protein Vpr arrests target cells in the G2 phase of the cell cycle and can induce apoptosis in T cells, peripheral blood lymphocytes, and fibroblasts (4). Once cells are actively producing virus proteins, the viral envelope protein gp160 is able to trigger apoptosis in cells as well (Gougeon and Debatin, 1999). Moreover, there is evidence that chronically HIV-infected cells can trigger programmed cell death in uninfected bystander T cells (Gougeon and Debatin, 1999). Finally, it has also been found that HIV-specific CD8 T cells have a weakened cytotoxic effect due to lower levels of perforin (Roeth, 2004). Lower levels of perforin make it less likely that the CTLs will be able to effectively penetrate and destroy infected CD4 cells that are productively producing new viruses.
References:
Janeway, Charles A, et al. Immunobiology. 6th ed. New York: Garland Science Publishing, 2005.
Finlay, B., McFadden, G. 2006. Anti-Immunology: Evasion of the Host Immune System by Bacterial and Viral Pathogens. Cell 124(4): 767-782.
Appay, V., et al. 2000. HIV-specific CD8+ T cells Produce Antiviral Cytokines but Are Impaired in Cytotoxic Function. Journal of Experimental Medicine 192(1): 63-76.
Gougeon, M., Debatin, K. 1999. Molecular control of programmed cell death in HIV infection. In: Dalgleish, A., Weiss, R., editors. HIV and the New Viruses. 2nd ed. San Diego: Academic Press, 1999.
Roeth, J. 2004. HIV-1 Nef Disrupts MHC-I trafficking by Recruiting AP-1 to the MHC-I Cytoplasmic Tails. Journal of Cell Biology 167(5): 903-913.