*This web page was produced as an assignment for an undergraduate course at Davidson College.*
Viral Description & Life Cycle
Hepatitis B virus (HBV) is a DNA virus of the hepadnaviridae family. The HBV virion, also called a Dane particle, is approximately 42 nm in diameter and consists of an inner protein core and an outer protein envelope. The outer envelope is composed of several proteins, known as hepatitis B surface proteins (HBs), which encase the nucleocapsid, or inner protein shell, made of hepatitis B core proteins (HBc) (Bruss and Ganem, 1991). The nucleocapsid is arranged into an icosahedral formation and contains the hepatitis B genome and at least one hepatitis B polymerase protein. The figure below shows a diagram of the DNA virus.
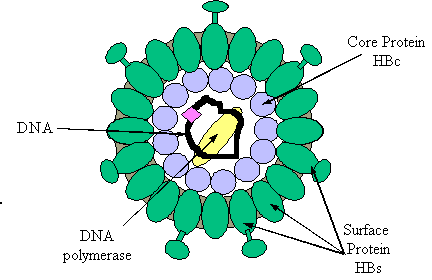
Figure 3. Image from http://www.hon.ch/Library/Theme/HepB/hbvirus.GIF
The viral genome is circular and is characterized by regions that are composed of single, double, and triple-stranded DNA. Within the genome are four partially overlapping open reading frames (ORFs) that encode seven different HBV genes, including but not limited to the hepatitis B surface antigen gene, also called the S gene, the hepatitis B core antigen gene, known as the C gene, the X gene, and the polymerase (P) gene (Ocama et. al., 2005). The figure below shows an image of the viral genome.
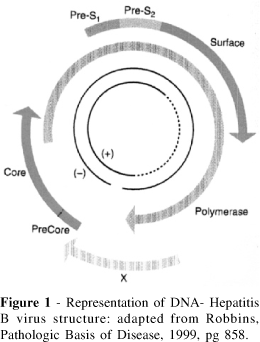
Figure 4. Image from http://www.scielo.br/img/revistas/rhc/v59n4/21391f1.gif
Unlike other envelope proteins, HBs span the lipid bilayer multiple times. There are three different size proteins based on the reading frame, yielding small, medium, and large HBs. The large hepatitis B surface antigen is believed to be most responsible for conducting viral-host cell attachment. A specific receptor, however, has yet to be identified. Another unique feature is that HBs can be secreted as subviral particles, lacking all other parts of the virion. Such particles are termed hepatitis B filaments and hepatitis B spheres, each of which has a diameter of 22nm. Increased levels can be found during acute infection. However, because these particles lack core proteins, polymerase, and viral DNA, they are non-infectious and do not appear to be advantageous to the virus (Bruss and Ganem, 1991).
Once the virus enters the body of a host, it infects liver cells. In doing so, the viral surface proteins fuse with the host cell membrane, allowing the core particle to enter the liver cell. Although early steps in viral entry are not clearly defined, the uncoated particle (or nucleocapsid) is transported into the nuclear membrane of the host cell. The viral DNA is then brought into the nucleus and is repaired to form a covalently closed-circular form, known as cccDNA. Viral DNA is not integrated into the host DNA like some viruses. Instead, once the DNA is recircularized, transcription of viral DNA and proteins begins and is controlled by four promoter elements and two enhancer elements. DNA polymerase then begins copying the DNA. The polymerase protein uses its unique shape to initiate reverse transcription and copy the DNA. Unlike retroviruses, hepadnaviruses bind polymerase proteins into a stem-loop formation, which are subsequently packaged by core proteins in the golgi and secreted via exocytosis into the blood stream, where it can contact other liver cells and continue replication (Rehermann and Michelina, 2005).
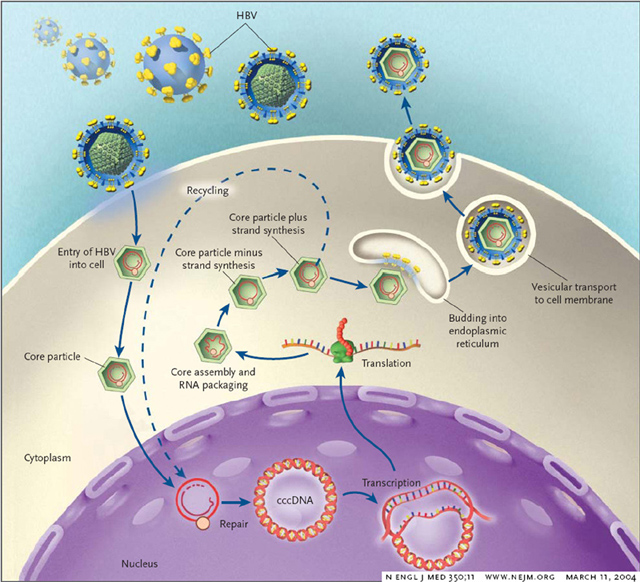
Figure 5. Image from http://mgl.snu.ac.kr/images/HBV%20life%20cycle.jpg
Ocama, P., Opio, C., & Lee, W. 2005. Hepatitis B virus infection: Current status. The American Journal of Medicine. 118(12):1413-1420.
Rehermann, B. and Nascimbeni, M. 2005. “Immunology of hepatitis B virus and hepatitis C virus infection.” Immunology 5: 215-229.
Christie Brough. Biology 307: Immunology. Dr. S. Sarafova. Davidson College. May 4, 2007.