**This webpage was produced as an assignment for an undergraduate course at Davidson College**
Cellular Immune Response
When HSV infects a cell, it controlled through the innate and adaptive immune responses (Khanna et al. 2004). Part of the adaptive immune response is the cell-mediated response in which naïve T cells are activated when they encounter their specific antigen by antigen-presenting cells. The T cell then differentiates and proliferates, creating a large number of CD4+ and CD8+ T cells specific for the antigen.
Primary Infection
A study by Gesser et al. (1994) demonstrated the importance of cell-mediated immunity. In severe combined immunodeficiency (SCID) mice (they have innate but no adaptive immunity) infected with HSV-1 in the ocular area, viral latency was established only in some and not all neurons. T cells were still needed for the mouse to recovery from the virus as SCID mice die of encephalitis (Gesser et al. 1994). The authors of the paper concluded that some part of the adaptive immune response is needed for the establishment of latency and control over the virus.
A follow up study looked at transferring T cells to SCID mice to determine if latency could be established in this fashion. The researchers found that when αβ TCR+ T cells were transferred to SCID mice, mice were able to stop HSV replication fully in the neuron and latency was established. In addition, they found that when CD8+ T cells were transferred, SCID mice experienced higher survival rates in primary infection compared to the negative control but they claimed that CD8+ T cell “engraftment seemed less needed for latency than that of CD4+ T cells” (Minagawa and Yanagi 2000). However, as this study is from 2000, the important role of CD8+ T cells will be discussed below.
More recent studies have elucidated the specific role of CD4+ and CD8+ T cells. In corneal HSV-1 infection, macrophages and γδ TCR T cells initially control viral replication, and this process is carried out by nitric oxide, IFN-γ, and TNF-alpha. After macrophages and γδ TCR T cells enter the infected area, CD4+ T cells and CD8+ T cells take over immune control (Khanna et al. 2004). The cytokine IL-12 induces CD4+ T cells to differentiate into Th1 cells, and these cells then activates CD8+ T cells (Cunningham et al. 2006).
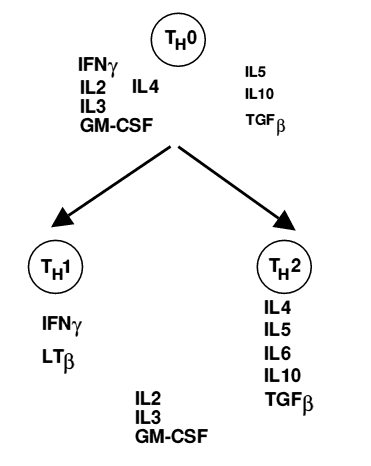
In particular, CD4+ T cells are the major cell type that invades the infected area and are detected 4-5 days after HSV-2 infection in the genital lymph nodes (Cunningham et al. 2006). CD4+ T cells play a role in halting viral replication and establishing latency (Khanna et al. 2004), and they mainly target viral gD and gB (Cunningham et al. 2006). CD4+ T cells also secrete IFN-γ (see Figure 2), which can repair MHC class I expression on infected cells (MHC class I expression is inhibited by the viral ICP47 protein). In addition, IFN-γ induces MHC class II expression on epithelial cells, which allows more CD4+ T cells to recognize the HSV antigen (Cunningham et al. 2006). Th1 and Th2 cells are differentiated CD4+ T cells, and HSV reactivation is prevented by Th1 cytokines (Aurelian 2004; see Figure 1). Cytokines produced by Th2 allow HSV to recur, and so effective treatments (especially with vaccines) will be directed toward stimulating a Th1 rather than a Th2 response (Aurelian 2004).
Figure 1. Cytokines produced by Th1 and Th2 cells. Image from http://www-immuno.path.cam.ac.uk/~immuno/part1/lec09/lec10_99.html
CD8+ T cells release IFN-γ and lytic granules, and these functions induce viral latency (Khanna et al. 2004). Bystricka and Russ (2005) found that CD8+ T cells invade the neuron approximately 7 days after HSV-1 infection, peak after 4 days, and then are present in the mouse for the lifetime of the animal. One of the CD8+ T cell targets is ICP27 (Cunningham et al. 2006). This information indicates that both subsets of T cells are present in clearing HSV infection and healing viral lesions (Rajcani and Durmanova 2006).
Figure 2. IFN-γ is an important cytokine in fighting HSV infection. Image from http://www.igb.fraunhofer.de/www/gf/Pharma/bilder/IFN-g-dimer_os_350.gif
Latency Period
Until recently, researchers did not understand the extent of the immune system’s involvement in HSV latency. Khanna et al. (2003) showed that a CD8+ T cell specific for gB(498-505) “blocked HSV-1 reactivation from latency in a dose-dependent, antigen specific and MHC-restricted fashion” when added to trigeminal ganglion cultures that lacked CD8+ T cells (Khanna et al. 2003). This study most importantly demonstrated that the immune system is active during HSV-1 latency.
CD8+ T cells maintain viral latency by monitoring infected trigeminal ganglion (Cunningham et al. 2006; Khanna et al. 2004). CD8+ T cells prevent HSV-1 reactivation through IFN-γ production. If added to cultures of neurons infected with latent HSV-1 that are early in starting reactivation, IFN-γ can block HSV-1 infection (Khanna et al. 2003). IFN-γ counters the effects of ICP0, an immediate early viral gene that reactivates HSV from latency, by inhibiting the transcription of ICP0 (Khanna et al. 2003).
Reactivation from the latency period can occur when CD8+ T cells are impaired, and one way in which CD8+ T cell function is damaged is through stress hormones (Khanna et al. 2004). Other factors that induce reactivation include ultraviolet light, heat, fever, hormonal changes, menstruation, and physical trauma to the neuron (Jenkins and Turner 1996). When reactivation occurs, the virus replicates and the nucleocapsid is created, but CD8+ T cells can detect epitopes of immediate early genes presented on MHC class I neuronal cell surfaces. The virion is then anterogradely transported down the axons (Khanna et al. 2004)
Click here to see an animation of Herpes Simplex Virus Infection
Davidson College Biology Department
Contact jehodge@davidson.edu with questions or comments.