
This web page was produced as an assignment for an undergraduate course at Davidson College
Zeta-chain-associated protein (ZAP-70)
Figure 1. Jmol image of ZAP-70 (RSCB Protein Databank)
Introduction.
Zeta-chain-associated protein (ZAP-70) is a 70kDa adaptor protein that acts
quickly after T cell activation to propagate signal. It is comprised of two
SH2 domains that bind phosphorylated ITAMs on CD3 and are joined by an interdomain
linking the two SH2 domains. This protein tyrosine kinase is present in both
CD4+ and CD8+ T cells and ultimately promotes gene transcription
and also has a role in apoptosis. Due to its importance in propagating the signal
from the TCR:CD3 complex, ZAP-70 is essential for a proper and complete T cell
response.
Structure.
ZAP-70 is comprised of two src homology 2 (SH2) domains connected by a central
interdomain linking region (Folmer et al., 2002). It is similar to
other SH2 domains of src family protein kinases in that it shares 35% of its
amino acid identity (Chan et al., 1992). SH2 domains comprise the phosphotyrosine
pocket and are named ZAP-C and ZAP-N for ZAP C-terminal SH2 domain and ZAP N-terminal
SH2 domain (Hatada et al., 1995).
ZAP-70 has two different conformations depending on whether or not it is bound to phosphorylated ITAMs or uncomplexed. The uncomplexed form has a different bond angle with respect to the linker region than does the ITAM-complexed form due to hydrophobic interactions between the ZAP-C and linker regions in the absence of phosphorylated ITAMs (Folmer et al., 2002; Figure 1). When complexed with ITAMs, the folding of the SH2 domains is similar to that of other SH2 domains on adaptor proteins in that they have a central ß-sheet flanked with two α helices; this yields a more stable molecule with a more definite conformation (Folmer et al., 2002; Figure 2).
ZAP-70 binds phosphorylated ITAMs via its SH2 domains (Figure 3). A myriad of contacts is made between the SH2 domains and the ζ-chain of CD3 on the TCR-complex with the most extensive contacts being between the phosphotyrosine and leucine. This type of interaction, mediated by the interdomain positioning the SH2 domains for binding, illustrates that the SH2 domains do not function independently of one another (Hetada et al., 1995).
Figure 1. Structure of ZAP-70 with interdomain region in red, ZAP-N in blue, and ZAP-C in green (Folmer et al., 2002)
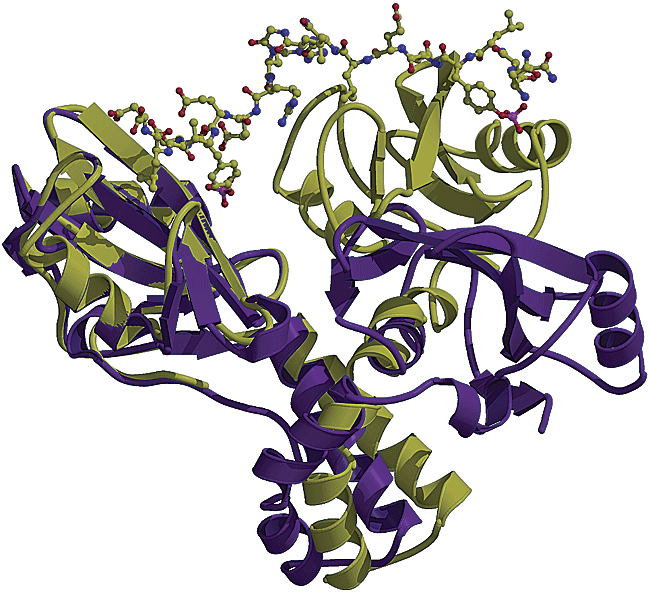

Figure 2. Left Frame: Conformation of ZAP-70 when bound to ITAM (tan) and when unbound (purple). Right Frame: Comparison of ITAM-bound (top) and unbound (bottom) forms of ZAP-70. Notice the binding pocket is completely closed in the unbound form (Folmer et al, 2002).
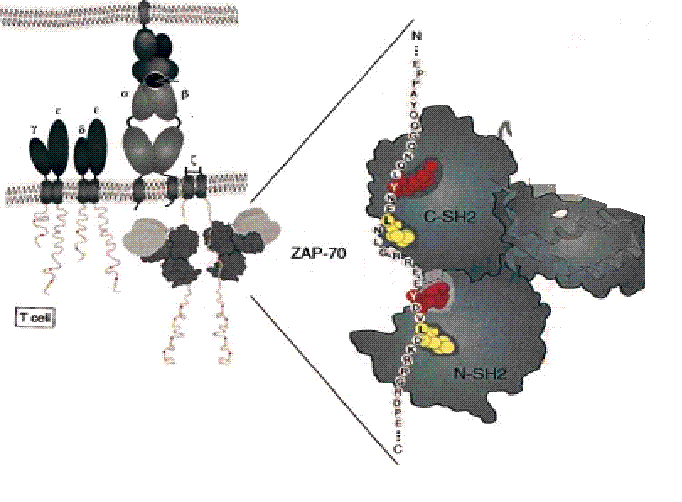
Figure 3. ZAP-70 binding to phosphorylated ITAMs (red) and leucine residues (yellow) of the ζ-chain via its SH2 domains (modified from Hatada et al, 1995)
Activation and Function.
When a T cell comes in contact with a peptide:MHC complex on an antigen presenting
cell, the protein kinase Lck or Fyn becomes activated via CD45. Lck or fyn then
up-regulates ZAP-70 by phosphorylating tyrosine 493 of ZAP-70 either by its
direct interaction with ZAP-70 or by allowing ZAP-70 to autophosphorylate by
causing structural rearrangements. These rearrangements allow the SH2 domains
to interact with the interdomain to autophosphorylate tyrosine 419 in the active
site. Catalytic efficiency is also seen via transphosphorylation with previously
phosphorylated ITAMs (Visco et al., 2000). These interactions occur
6-15 seconds after TCR clustering (Bunnell et al., 2002).
ZAP-70 activates two proteins: LAT (linker of activation in T cells) and SLP-76.
LAT is an adaptor protein that performs signal transduction in a lipid-raft
mediated function. It is phosphorylated by activated ZAP-70 to create docking
sites for other SH2 domains, including phospholipase C-γ (PLC-γ) and SLP-76 (Tanimura
et al., 2003). The activation of PLC-γ initiates the exhaustedly
studied pathway, beginning with cleavage of PIP2 and the release of calcium,
ultimately activating transcription factors NFκB and NFAT. SLP-76 plays
a crucial role in activating the MAP kinase signaling pathway used to transcribe
the AP-1 transcription factor. LAT, in conjunction with Grb2 (another adaptor
protein) allows SLP-76 to activate SOS, a GEF involved in the pathway, which
stimulates the small G-protein Ras to ultimately up-regulate the transcription
factor Elk to produce Fos, half of the AP-1 transcription factor (Kumar et
al., 2002). SLP-76 and LAT also work to activate another GEF, Vav, which
also initiates a MAP kinase cascade to produce Jun, the other half of the AP-1
transcription factor (Janeway et al., 2005).
New research also shows ZAP-70 has a role in apoptosis. TCR-mediated apoptosis takes place via the ζ-chain that ZAP-70 associates with. Although the exact mechanism is unknown, ZAP-70 deficient T cells did not express FasL and undergo apoptosis (Zhong et al., 2004).
Deficiencies and Clinical Applications.
Severe combined immunodeficiency (SCID) occurs when there is an absence or deficiency
in ZAP-70. This is characterized by low levels of CD8+ T cells in
the periphery and TCR deficient signaling. The absence of CD8+ T
cells illustrates that ZAP-70 is essential for CD8 development, however, the
presence of CD4+ T cells demonstrates a different possible pathway
is available involving a similar adaptor protein, Syk. Syk is normally found
in high concentrations in B-cells, but very low and often untraceable amounts
in T cells. The Syk pathway does not interact with ITAM domains, however allows
for an incomplete activation of peripheral T cells in the immune response, leading
to an immunocompromised system (Hivroz and Fischer, 1994; Figure 4). For example,
patients with SCID may not be able to induce isotype switching and produce specific
antibodies. A study by Toyabe et al (2001) shows ZAP-70 deficient patients
are able to express CD40L and secrete IL-4 to induce isotype switching as a
result of increased expression and activation of Syk.
The FDA has not approved any drugs which act specifically on ZAP-70. However, SH2 inhibitors are being researched as possible immunosuppressants. A series of highly selective 1,2,4-oxadiazole analogues have been studied which show specificity for only the ZAP-70 SH2 domains by mimicking ζ-chain domains (Vu et al., 1999). This discovery is exciting because with only the inhibition of ZAP-70, an incomplete immune response can still be formed via the Syk pathway, but, theoretically, not large enough to cause rejection or other immune problems.
More specifically, a point mutation in ZAP-70 has been shown to cause Rheumatoid Arthritis in mice. Rheumatoid arthritis is an autoimmune disease that affects 1% of the world population characterized by symmetrical swelling in the joints. This point mutation causes a change in sensitivity for developing thymocytes, causing the positive selection of autoimmune T cells that should be negatively selected against. Sakaguchi et al (2003) proposes the signals that occur during positive and negative selection must involve ZAP-70 and this mutation causes an increase in avidity for self-peptide:MHC complexes to compensate for the impaired signal. They further theorize that other autoimmune diseases do not appear because most autoimmune diseases destroy their target cells, while Rheumatoid Arthritis stimulates synoviocytes to proliferate and secrete inflammatory cytokines (IL-1, IL-6, TNF-α) that destroy cartilage and bone (Sakaguchi et al., 2003).
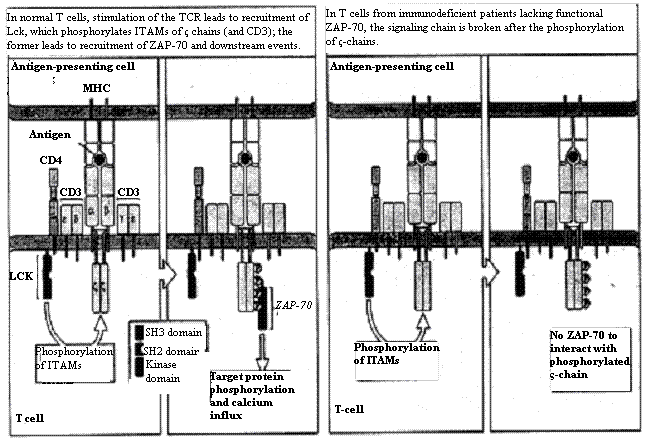
Figure 4. Normal signaling pathway (left) and ZAP-70 deficient pathway (right). ZAP-70 deficient cells still demonstrate some immune response via Syk, however, Syk does not interact with ζ-chains (modified from Hivroz and Fischer, 1994).
References.
Bunnell, SC, DI Hong, JR Kardon, T Yamazaki, CJ McGlade, VA Barr, LE Samelson.
(2002). T cell receptor litigation induces the formation of dynamically regulated
signaling assemblies. Journal of Cell Biology 158: 1263-1275.
Chan, AC, M Iwashima, CW Turck, A Weiss. (1992). ZAP-70: a 70kd protein-tyrosine kinase that associates with the TCR ζ chain. Cell 71: 649-662.
Folmer, RHA, S Geschwinder, Y Xue. (2002). Crystal structure and NMR studies of the apo SH2 domains of ZAP-70: two bikes rather than a tandem. Biochemistry 41: 14176-14184.
Hatada, M.H., X. Lu, E.R. Laird, J. Green, J.P. Morgenstern, M. Lou, C.S. Marr, T.B. Phillips, M.K. Ram, and K. Theriault. (1995). Molecular basis for interaction of the protein tyrosine kinase ZAP-70 with the T cell receptor. Nature 377: 32-38.
Hivroz, C, A Fischer. (1994). Multiple roles for ZAP-70. Current Biology 4: 731-733.
Janeway, CA, P Travers, M Walport, MJ Shlomchik. (2005). The development of mature lymphocyte receptor repertoires. In: Immunobiology (Lawrence, E). Garland Science Publishing, 218-221
Kumar, L, V Pivniouk, MA de la Fuene, D Laouini, RS Geha. (2002). Differential role of SLP-76 domains in T cell development and function. Proceedings of the National Academy of Sciences of the United States of America 99: 884-889.
Sakaguchi, N, T Takahashi, H Hata, T Nomura, T Tagami, S Yamazaki, T Sakihama, T Matsutani, I Negishi, S Nakatsuru, S Sakaguchi. (2003). Alterd thymic T cell selection due to a mutation of the ZAP-70 gene causes autoimmune arthrities in mice. Nature 426: 454-460.
Tanimura, N, M Nagafuku, Y Minaki, Y Umeda, F Hayashi, J Sakakura, A Kato, DR Liddicoat, M Ogata, T Hamaoka, A Kosugi. (2003). Dynamic changes in the mobility of LAT in aggregated lipid rafts upon T cell activation. Journal of Cell Biology 160: 125-135.
Toyabe, S, A Watanabe, W Harada, T Karasawa, M Uchiyama. (2001). Specific immunoglobulin E responses in ZAP-70-deficient patients are mediated by Syk-dependent T cell receptor signaling. Immunology 103: 164-171.
Visco, C, G Magistrelli, R Boscotti, R Perego, L Rusconi, S Toma, M Zamai, O Acuto, A Isacchi. (2000). Activation of ZAP-70 tyrosine kinase due to structural rearrangement induced by tyrosine phosphorylation and/or ITAM binding. Biochemistry 39: 2784-2791.
Vu, C, EG Corpuz, TJ Merry, SG Pradeepan, C Bartlett, RS Bohacek, MC Botfield, CJ Eyermann, BA Lynch, IA MacNeil, MK Ram, MR van Schravendijk, S Violette, TK Sawyer. (1999). Discovery of Potent and Selective SH2 Inhibitors of the Tyrosine Kinase ZAP-70. J. Med. Chem. 42: 4088-4098.
Zhong, L, CH Wu, WH Lee, CP Liu. (2004). Zeta-Associated protein of 70 kDa (ZAP-70),
but not Syk, tyrosine kinase can mediate apoptosis of T cells through the Fas/Fas
ligand, caspase-8 and caspase-3 pathways. Journal of Immunology 172:
1472-1482.
Created by: Joseph
Scognamiglio ![]()
Return to: Immunology Homepage
Return to: Davidson College Homepage