This page has been created as part of an undergraduate course
at Davidson College
Transformation
WHAT IS TRANSFORMATION?
By definition, transformation is the bacterial mechanism for
the transfer of genetic material in which free DNA of one genotype is taken
in through the cell surface of bacteria of another genotype and is incorporated
into the recipient cell chromosome. In eukaryotic cells, the term transformation
refers to the conversion of normal eukaryotic cells into a cancer-like state
(Purves et al., 1998). The term transfection is used to describe uptake, incorporation,
and expression of foreign DNA into eukaryotic cells (Unknown, 2003).
HISTORY OF TRANSFORMATION
Griffith was the first to observe transformation. In his classic experiment,
Griffith transformed nonvirulent pneumococcus to virulence with DNA (Purves
et al., 1998).
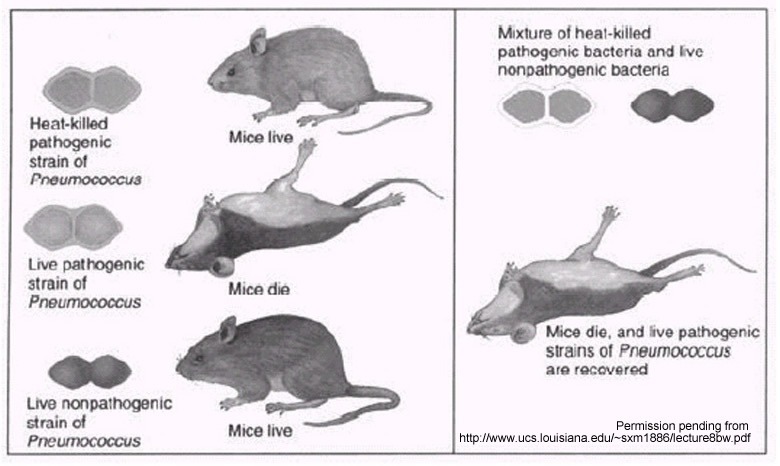
Fig. 1. Griffith's experiment. This figure illustrates
that when mice were infected with heat-killed pathogenic (S) strain of Pneumococcus
the mice lived, as opposed to being infected with the live pathogenic strain,
which caused death. Infection with the live nonpathogenic (R) strain did not
cause the mice to die. Mixing the heat-killed pathogenic strain and the live
nonpathogenic strain, however, did cause the mice to die. Live pathogenic strains
of Pneumococcus were recovered from the mice that died of the mixed
infection.
Griffith showed that a particular factor in the virulent S strain could transform
the nonvirulent R strain of pneumococcus into a lethal form, even when the S
strain had been killed after exposure to high temperatures. We now know that
DNA had escaped from the dead S cells and that the living R cells had then picked
up the DNA within the body of a living mouse (Purves et al., 1998).
HOW DOES IT WORK?
Not all bacteria take up free-floating DNA in the environment. The genera that
generally exhibit transformation include: Bacillus, Streptococcus, Azotobacter,
Haemophilus, Neisseria, and Thermus. The recipient cells must be competent (able
to transform). Competence is a phenotype conferred by one or more proteins.
It has been shown that competence occurs late in the exponential phase of bacterial
growth. The duration of competence varies from a few minutes in Streptococcus
to hours in Bacillus (Krawiec, 2002).
Dubnau and Provvedi (2000) described the proteins involved in the steps of
B. subtilis transformation as shown in table 1 below.
Table 1. B. subtilis competence proteins
| CLASS |
PROTEIN |
LOCATION |
PROPOSED ROLE |
| I |
ComEA |
Bitopic integral membrane protein, with C-terminus outside |
DNA receptor and presentation of DNA to transport machinery |
| II (PSTC) |
ComC |
Polytopic integral membrane protein (inferred from sequence) |
Processing protease |
| II (PSTC) |
ComGA |
Peripheral membrane protein on inner face of membrane |
Access of DNA across wall |
| II (PSTC) |
ComGB, ComGF |
Integral membrane proteins (inferred from sequence) |
Access of DNA across wall |
| II (PSTC) |
ComGC, ComGD, ComGE, ComGG |
Integral membrane proteins, translocated to wall when processed by ComC |
Access of DNA across wall |
| III |
ComEC |
Polytopic integral membrane protein (inferred from sequence) |
DNA transport: (channel?) |
| IV |
ComFA |
Integral membrane protein, exposed on inner face of membrane |
DNA translocator |
Table courtesy of Dave Dubnau
In essence, double stranded DNA binds to the surface of a competent cell and
is cleaved into fragments of about 15 kb. The double stranded DNA, which is
still external to the cell, is then separated into single stranded DNA. One
fragment of the single stranded DNA is degraded, while the other is transported
across the membrane and into the cell (Dubnau et al., 2000). Upon entry into
the cell, a single strand of the foreign DNA is incorporated into the recipient
cell via recombination (Krawiec, 2002).
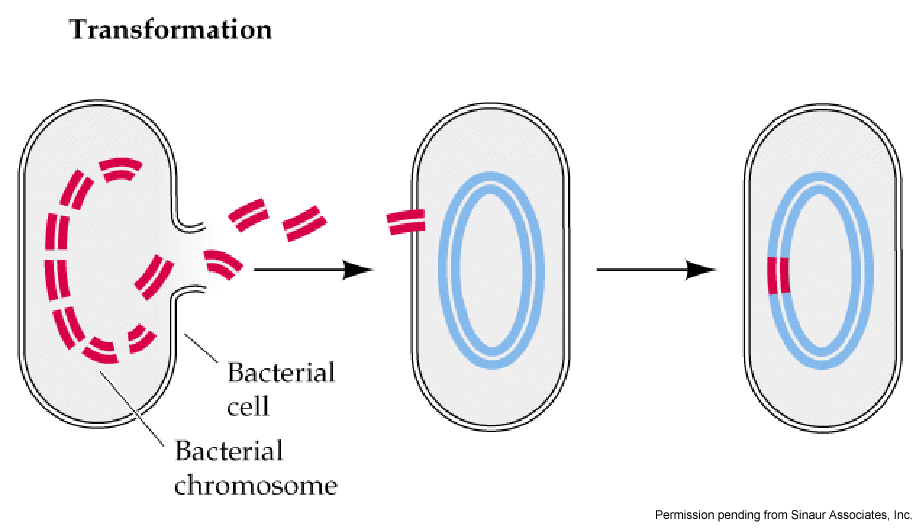
Fig. 2. Illustration of bacterial transformation.
DNA from dead cells gets cut into fragments and exits the cell. The free floating
DNA can then be picked up by competent cells. The exogenous DNA is incorporated
into the host cell's chromosome via recombination.
IN THE LAB
Transformation is used in the lab in order to assess linkage. Extraction of
the donor DNA causes inevitable breakage. The relative map distance is determined
according to the percentage of two genes being on the same transforming segment
(double transformation). For example, two genes that are close to each other
on the chromosome will have a high occurrence of double transformation and a
low occurrence of being carried on separate transforming segments (Griffiths
et al., 2000).
In performing linkage experiments, the major problem of inducing transformation
must be over come. The exterior surface of the plasma membrane is negatively
charged due to the phospholipid head groups. This results in repulsion of the
negatively charged DNA. The charge repulsion can be temporarily fixed by neutralizing
the DNA with calcium salts. The plasma membrane is permeable to DNA when the
charge has been neutralized. This technique is used on both prokaryotic and
eukaryotic cells being transformed. In plants and fungi, however, the cell wall
must first be removed with enzymes, which results in protoplasts (Griffiths
et al., 2000).
FOR
MORE ON TRANSFORMATION.....
REFERENCES
Dubnau, D., Provvedi, R. (2000). Internalizing DNA. Research in Microbiology,
151: 475-480.
Griffiths, A.J.F., Gelbart, W.M., Miller, J.H., Lewontin, R.C. (2000). Modern
Genetic Analysis. New York: W.H. Freeman and Company.
Krawiec, S.S. (2002). Prokaryotic Genetics. Lehigh University. 12 Feb 2003.
www.lehigh.edu/~sk08/courses/Bios324/Module9.htm.
Purves, W.K., Orians, G.H., Heller, H.C., Sadava, D. (1998). Life. Sunderland,
Massachusetts: Sinauer Associates, Inc.
Unknown. Recombinant Protein Production in Eukaryotic Cells. 12 Feb 2003. http://classweb.gmu/archriste/385-Ch07appt/385-Ch07a-PwrPt.
Please send any questions or comments to: Monica
Siegenthaler
Molecular Biology at Davidson
College
Back to my molecular biology home page
Spring 2003
