CAT8 & YMR279C: My Favorite Yeast GenesPart II: Analysis with Expression Profiles |
The free-access ethos pervading microarray technology and the massive jungle of experimental data that first sprouted in its wake 10 years ago are at the heart of genomics. Microarrays are fundamentally good at sampling the expression patterns of genes in a highly parallel manner, making experiments based on whole-genome surveys not just possible, but practical and economical. Public databases have been deluged with data ever since, and it is actually becoming more expensive and difficult to successfully organize all the data logically than physically. Join us as we set sail across that sea of data in Part II, fishing for more clues to the function and ontology of YMR279C & CAT8.
In part I we investigated the function of the S. cervisiae genes CAT8 and YMR279C using existing literature0, ontology & sequence databases such as the Saccharomyces Genome Database, and a variety of in silico methods such as BLAST to identify other genes with similar sequences. By assuming that function follows form (in the microscopic world, at least), we could infer some of the functions of our genes from any results that had already been annotated. This strategy can be extended for use with expression patterns: if gene A is well understood and gene B unknown, but both share a great deal of similarity in expression, then perhaps both participate in the same cellular process. This reasoning process is generally called "guilt by association" and increases in power with the similarity and number of expression patterns. Clustering functionally-similar genes based on expression profile can include genes that a BLAST method would ignore, if they were functionally related but composed of totally different sequences.
On the other hand, we could forgo the other genes completely and just look at how the expression levels of those genes in which we are interested react to a comprehensive battery of tests like heat/cold shocks, changes in nutrition, starvation, drawing conclusions about their functions based on their individual response. But the technology to look at single-gene expression has been around for some time now, and the real strength of microarray experiments is in their ability to reveal trends and patterns between many groups of many genes.
 |
CAT8 |
|
CAT8's Molecular Function is officially annotated as having "Specific RNA polymerase II transcription factor activity" and it's Biological Process being involved with "Positive regulation of gluconeogenesis." Before attempting to devine functional information for YMR279C (of which we know very little) from expression data, let's test out the technique on CAT8.
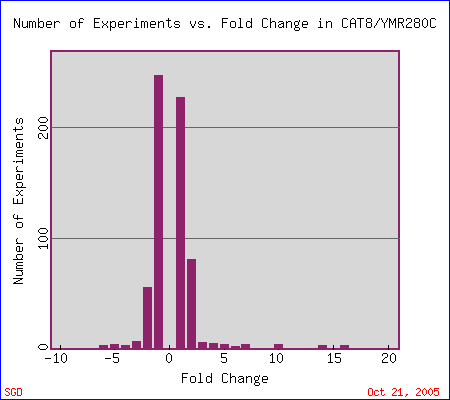
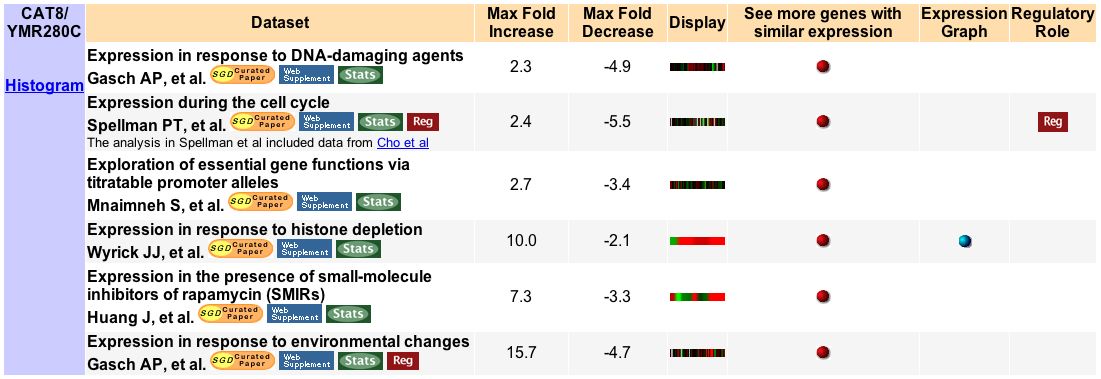
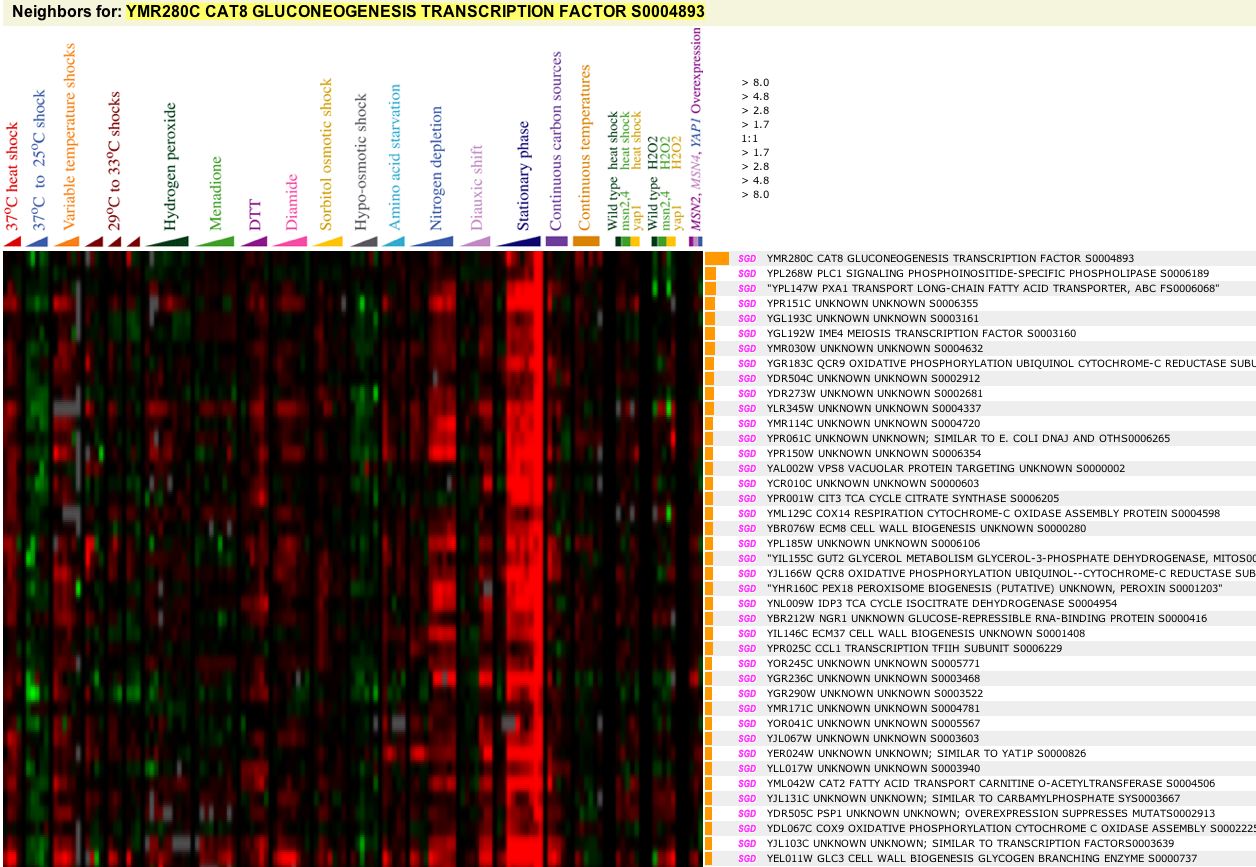
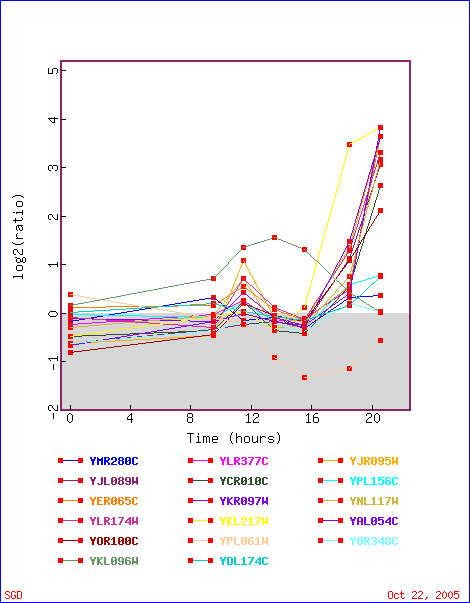
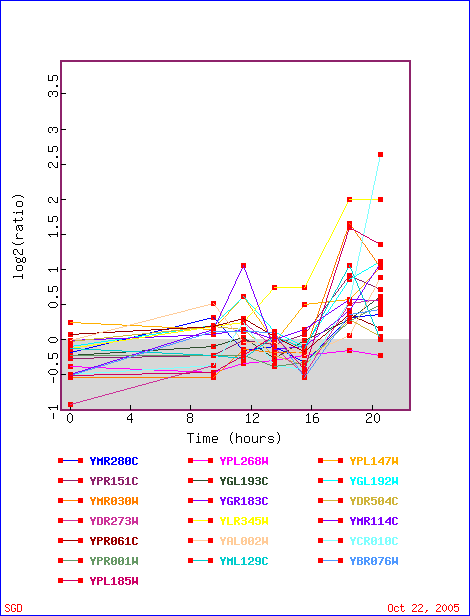
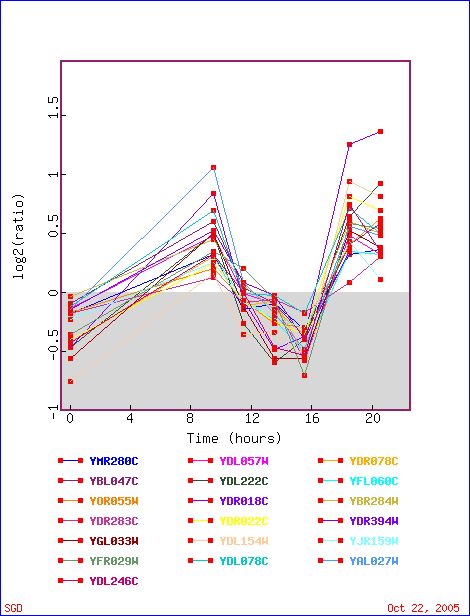
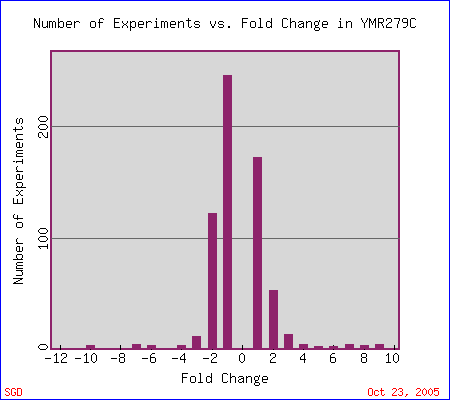
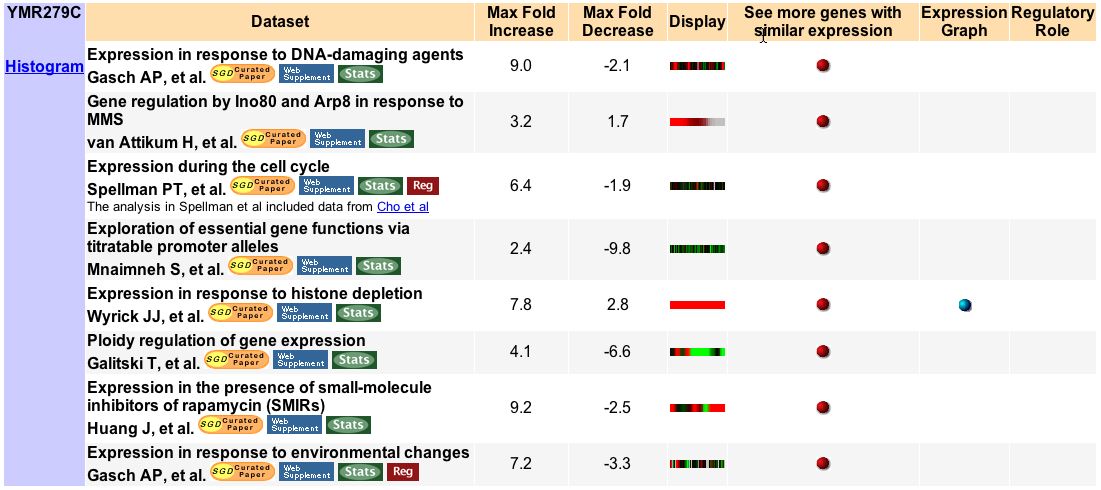
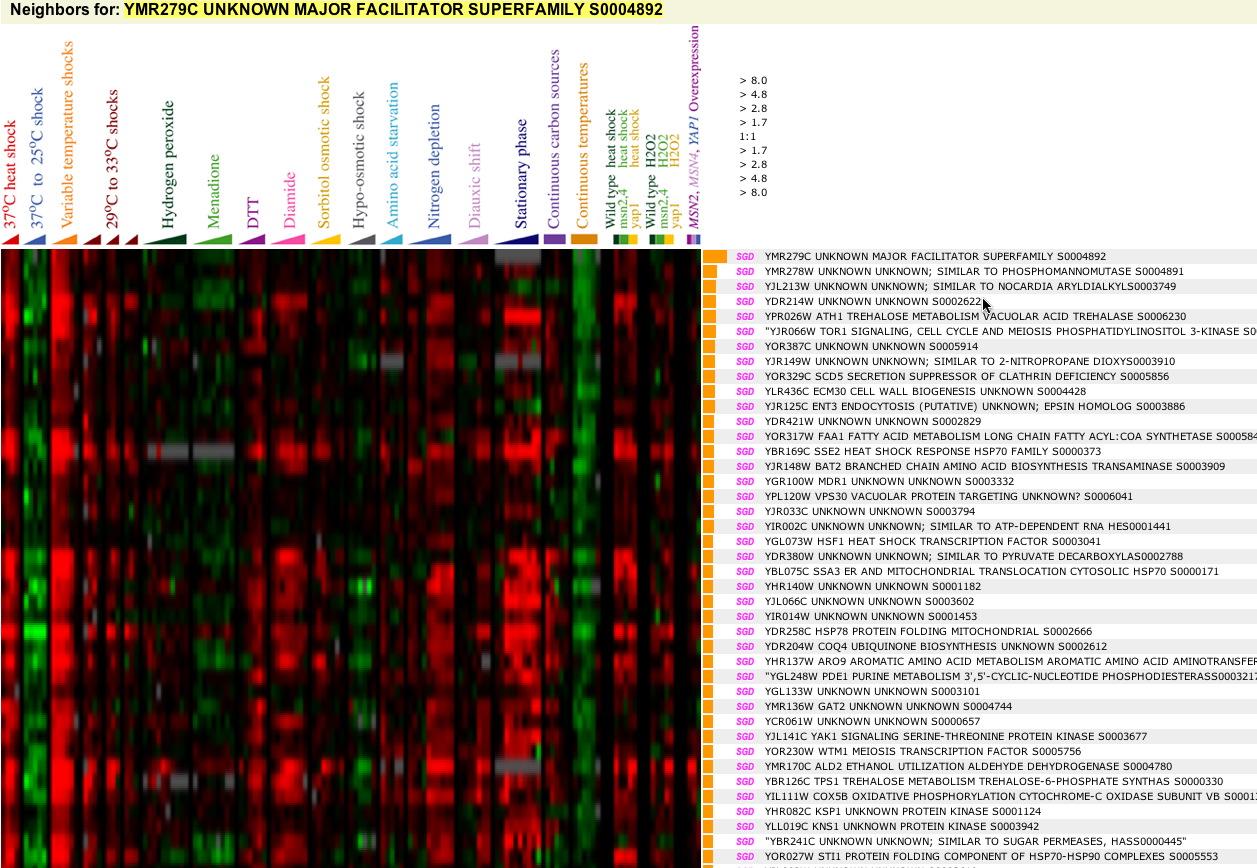
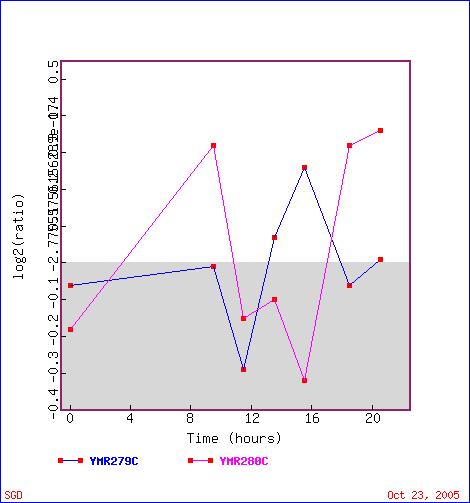
First stop: the Saccharomyces Genome Database, host of the Expression Connection. fig. 2 presents an overview of CAT8's fold change amongst all of the microarray experiments in the database, revealing that it is usually only subtly induced or repressed. This is consistent with the notion that it is an early player in the pathway that ultimately causes the diauxic shift - it only needs to interact with a limited number of other proteins & binding sites. fig. 3 presents several of the less common experiments in which CAT8 experiences a 3+fold change in expression. We can use this perspective to help decide what datasets to evaulate first, and then begin exploring them. However, just because our gene underwent a large fold change doesn't guarantee the usefulness of the dataset - in some cases it isn't clear what aspect of the experimental conditions caused that change, and in other cases, especially those testing many different experimental conditions, no other genes have a similar pattern of expression, at least by the metrics used on the Expression Connection website. This is the case for several of the datasets listed below, including "Expression in response to environmental changes" by Gasch et al (2000). Exploring the data directly from the supplementary website is more informative (fig. 4). Notice the high amounts of induction in the "stationary phase" column. This is consistent with CAT8's role in regulating the diauxic shift.
{kind=link}