The Role of Cell-Mediated Immunity
The cellular immune response involves the adaptive immune response in which antigen-specific T cells play the main role in controlling and eliminating the infection. This is a type of adaptive immunity that can't be transferred to a naive recipient with the serum antibody. (Janeway et al., 2005) When R. rickettsii invades a host cell, it kills cells in absence of immune system effectors and is able to manipulate the host’s immune system to for its own benefit. (Walker et al., 2003)
There is a great deal of research supporting the primary role of the cell-mediated immune response for successful clearance of Rickettsia rickettsii from an infected host. Many studies have shown that the passive transfer of T cells from an immune host will sufficiently protect a naïve animal. There are a few speculative mechanisms as to how to obtain rickettsial immunity, many of which been examined through assays of cell-mediated immunity. Examples of this include delayed hypersensitivity and proliferation of T cells in the presence of a specific antigen. It is currently hypothesized that the cell-mediated immune response is necessary for both clearing an infection from a host cell and for immunity to future infections. Existing evidence supports that antigen-responsive T cells, specifically T helper cells, play this critical role in immunity through cellular effector mechanisms, as well as the production of T-cell dependent antibodies. (Gage et al., 1992)
Additional evidence also suggests that other nonpathogenic rickettsial species are able to induce cross-reactive cellular and humoral immune responses to R.rickettsii infections. A study done by Gage and Jerrells in 1992 showed that R.rhipicephali, a low virulent pathogen not closely related to R.rickettsii, responded to the lymphocytes of R.rickettsii as good or better than to their own immunizing antigen. This cross-protective study performed in guinea pigs could help discover the proteins responsible for this phenomenon and determine how protective immunity could be utilized to prevent the infection. The effectiveness of the cross-protective response in this experiment depended on the proliferation of antigen-responsive T lymphocytes to R. rickettsii in the host. Such cross-reactivity is speculated to be the result of similar levels of cross-reactive T-cell epitopes recognized by antigen-specific T lymphocytes. One of these specific antigens was rickettsial LPS, but this response may have been due to the presence of a chronic infection. In addition, this study found that the rOmpB antigen is important in protective immunity against R.rickettsii. This protein has also been found to be a major factor in the stimulation of antigen-responsive helper T lymphocytes. Specific mechanisms of this cross-reactivity are still not known.
Gimenez stain of tick hemolymph cells infected with R. rickettsii. 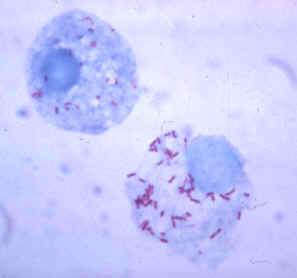
Photo and text courtesy of the CDC - http://www.cdc.gov/ncidod/dvrd/rmsf/Organism.htm
Mechanism of Cell-mediated Immune Response
When R.rickettsii is detected within the cell, the adaptive immune system triggers the activation of cytotoxic CD8 T cells to eliminate the infected cells through apoptosis. If there is an unusually large target of infected cells, they can be killed by necrosis before the T cells arrive. In addition, the NFkB pathway is often activated in order increase the response at the concentrated areas so that the increased production of cytokines and chemokines can result in a greater recruitment of cells to the site of infection. (Walker et al., 2003) There are a variety of specific cytokines and chemokines that are secreted in response to the activation of the intracellular signaling cascades that result in differential gene expression.
Studies have shown the Rickettsia rickettsii infections results in the production of specific immune effector molecules through a variety of intracellular signaling mechanisms. Current research has documented the rapid, dose dependent activation of the p38 MAP kinase pathway due to increased phosphorylation and enzymatic activity, but not of the JNK pathway. This unique selectivity of the activation of MAP kinase but not JNK shows strict regulation through a stress-induced mechanism. Oxidants, antioxidants, and other determinants of the intracellular redox state can greatly affect the intracellular signaling pathways. Measurements of the reactive oxygen species, along with some other damaging mechanisms, play a critical role in oxidative stress and adaptive antioxidant mechanism that directly influence the pathogenicity of R.rickettsii. (Rydkina et al., 2005) Some additional damaging mechanisms could include the production of nitric oxide, reactive oxygen species, and limiting of availability of tryptophan through the degradation by indoleamine-2,3-dioxygenase. (Walker et al., 2003) There is strong evidence to support that there is a direct connection between p38 and the NFkB pathway as well, as well as p38's involvement in the regulation of cytokine production. (Rydkina et al., 2005)
Cytokines are proteins that are made by cells that affect the behavior of other cells. (Janeway et al., 2005) These cytokines produced by R. rickettsii are activated intracellularly by rickettsial activity in various combinations by three different mechanisms. Once cytokine activation is complete, human cells will secrete IFN-gamma, TNF-alpha, IL-1Beta, and RANTES into the blood stream. (Clifton et al., 2005) Several studies have even shown that the T-lymphocyte associated immune response is mediated by interferon gamma. There are still a wide range of studies being conducted to attempt to explain how this process occurs. (Williams et al., 1986)
Chemokines are small chemoattractant proteins that stimulate the migration and activation of cells. When endothelial cells are infected with Rickettsia rickettsii, within three hours after infection, the chemokines interleukin-8 (IL-8) and monocyte chemoattractant protein-1 (MCP-1) are produced. These proteins are essential in the activation and recruitment of neutrophils and monocytes to the sites of inflammation. This specific chemokine response was studied using a peptide-aldehyde compound MG132 that prevents the degradation of inhibitor IkappaBalpha and SN-50 that prevented signaling molecule NF-kappaB from entering the nucleus and inducing gene transcription (this inhibits the response). The secretion of IL-8 can be down-regulated by T24 cells expressing a super-repressor mutant of IkappaBalpha. Though many experiments are still being performed, the chemokine response early during the infection depends on the direct activation of the NF-kappaB signaling pathway. (Clifton et al., 2005) This type of infection is thought to be one possible mechanism that provides long-lasting immunity against reinfection. (CDC, 2005)