Infection/Tranmission
The life cycle of Rickettsia rickettsii involves a sequence of events including the transmission of the disease, entry at a particular anatomic site, the spreading of the disease, evasion of host defenses, growth within a specific location in the body, and the mechanism of injury caused to the cells, tissues, organs, and the rest of the host's body. ( Walker, 2003)
There are two main types of host cell cycles for R. rickettsii. They have natural cycles with parasitic systems that involve many wild mammals, tick vectors, and long term reservoirs of rickettsiae. Secondly, they also have domestic or anthropogenic cycles that involve domestic animals, especially dogs. People are an accidental vector in the circulation and maintenance of R. rickettsii. These pathogens will mutate quickly and have shown a broad degree of severity in humans especially. (Rudakov et al., 2003)
R.rickettsii often infects the tick family Ixodidae (hard ticks). These ticks have four stages in their life cycle: egg, larva, nymph, and adult. Ticks are infected with R. rickettsii while feeding on blood from the host in the larval, nymphal, or adult stage. Once they have the pathogen, they can remain infected for life. After an immature tick develops into the next stage, this pathogen can be transmitted to a secondary host. R.rickettsii is transmitted to a vertebrate host through saliva while the tick is feeding. In addition, male ticks can transfer the pathogen to female ticks through bodily fluids or spermatozoa during mating, and female ticks can transmit the bacteria to her eggs through transovarial transmission. The two most common species of tick vectors in the United States are the American Dog Tick (Dermacentor variabilis) and the Rocky Mountain Wood Tick (Dermacentor andersoni). These ticks feed on many vertebrates such as dogs, medium-sized mammals, and small rodents. (CDC, 2005)


Rocky Mountain Wood Tick
Photo courtesy of the CDC - http://www.cdc.gov/ncidod/dvrd/rmsf/Natural_Hx.htm
R. rickettsii are obligate intracellular parasites. They are often spread through arthropod vectors beyond just ticks, including lice, fleas, and mites. Once they are within their arthropod vectors, the bacteria multiply in the epithelium of the intestinal tract. They are excreted through feces, but sometimes will obtain access to the salivary glands. They are transmitted to humans through arthropod saliva often by a bite. They multiply through transverse binary fission. In poor conditions, they stop dividing and grow into long filamentous forms. In these circumstances, they undergo many excessive, rapid divisions into the typical short rod forms. Following division, they are free to move throughout the cytoplasm in the cell. (CDC, 2005)
Host Cell Entry
In order to begin enter a host cell, the R. rickettsii attach to a protein-dependent receptor on the cell membrane of the host. (Walker et al., 2003) The cell wall of R. rickettsii is composed of the outer membrane, peptidoglycan, and cytoplasmic membrane. (Williams et al., 1986) With the aid of outer membrane protein A (OmpA), the adhesion of this molecule to the host cell induces local cytoskeletal rearrangements with the cell, which results in their entry into the cell. (Walker et al., 2003) The specific mechanism by which the pathogen actually enters the cell is not completely understood. Both the host cell and the microorganism must participate in order for R.rickettsii to successfully internalize. It is also suggested, based on evidence found in R. prowazekii, that these parasites enter through a mechanism that involves phospholipase A activity because host cells exposed to large numbers of rickettsial organisms release considerable amounts of lysophophatides and free fatty acids into the cell culture medium. In addition, one study by Silverman et al. examined whether the phospholipase involved in the penetration and lysis of erythrocytes originates from the rickettsiae itself or if it is a host cell enzyme that becomes activated upon contact of the pathogen with the plasma membrane. Their experiments supported that the entry of these cells into eukaryotic host cells is likely mediated by a rickettsial phospholipase. This study provided insight into the role of phospholipase in R.rickettsii’s entry into a cell that took into account the specificity of the inhibition of rickettsial entry and the toxicity of specific inhibitors of phospholipase activity on the host cell and organism itself. Both an irreversible inhibitor of phospholipase A2 and antiserum to phospholipase were effective methods for blocking the internalization of R.rickettsii, the antibodies in antiserum lasting for an extended time due to their stability. (Silverman et al., 1992)
There are many possible methods of transmission in which R. rickettsii could enter a host cell. These tick-bourne microorganisms are transmitted into a host cell transoverially or transstadially. One possible explanation for the difference in severity of Rocky Mountain Spotted Fever cases in humans may be the result of the different stages of the vector’s metamorphosis (and therefore different physical structure) upon infection. There is speculation of antigenic mimicry resulting from the simultaneous existence of larvae and nymphs in the same carrier. (Rudakov et al., 2003) Most commonly, however, this pathogen is transmitted to a human through an arthropod vector, most often a tick. (Walker et al., 2003) The main vector species are the ticks of genera Dermacentor, Rhipicephalus, Haemophysalis, Ixodes, Amlyomma, and some insects. (Rudakov et al., 2003) R. rickettsii can enter the human host through salivary inoculation by feeding tick, deposition of feces, and aerosol. It can enter through the skin, mucous membranes, and the lungs. It spreads through the lymphatic vessels from where it entered to lymph nodes in the surrounding regions, as well as through the bloodstream to all of the organs of the body. It often targets the epithelium, especially focusing the infection in the brain and the lungs. (Walker et al., 2003)
The most recent hypothesis for the entry of R.rickettsii into its host cell involves the rickettsiae binding to a specific protein on the membrane surface, inducing phagocytosis, and escaping from the phagosome into the cytosol. This mechanism is thought to parallel a model similar to Rickettsia conorii, where OmpB, the most abundant surface protein in rickettsiae, engages Ku70, a protein that spans the host cell membrane. Martinez et al. found that Ku70 migrates to the host membrane when rickettsiae are present, and induce downstream signaling cascades that may eventually result in the initiation of endocytosis. With the binding of Ku70 to OmpB, the enzyme ubiquitin ligase is recruited to the binding site to adding an ubiquitin moiety to Ku70. Research studies are still investigating therapeutic treatment methods involving Ku70 and ubiquitination. The current evidence suggests that there are one or more alternative receptors for rickettsial attachment proteins, in addition to one or more attachment proteins. The previous hypothesis of this group that suggests signal transduction pathways cause ubiquitinated Ku70 to result in cytoskeletal rearrangements to trigger the phagocytosis of rickettsiae. Revolutionary treatments for R.rickettsii could target the escape of bacteria from phagosomes or the cell-to-cell spread of rickettsiae through the release of actin into the host cell. These new treatments could produce antibodies to OmpB, inhibition of the functions of host-cell proteins that are necessary for rickettsiae to survive, and numerous other possibilities associated with this mechanism. ( Walker, 2006)
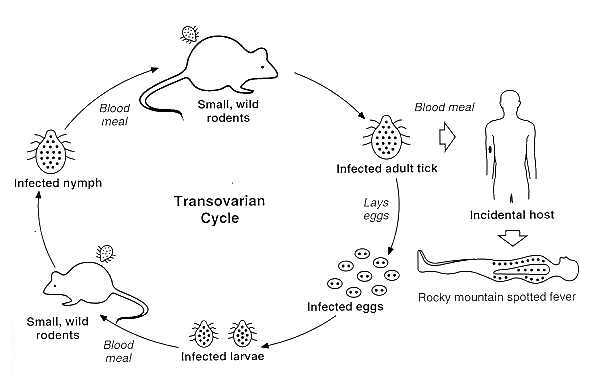
The above figure shows the transovarian passage of R rickettsii in the tick vector. Horizontal transmission (i.e., acquisition of the bacteria by uninfected ticks feeding on infected animals) occurs less often and is not shown. Humans become incidental hosts after being bitten by an infected adult tick.
Photo courtesy of David H. Walker - http://gsbs.utmb.edu/microbook/ch038.htm
Damage following Infection
Once the bacteria have entered the cell, it allows them to be taken up through nonprofessional phagocytes through a mechanism that requires rickettsial metabolic activity. Once into the phagosomes, the bacteria escape into the cytosol by lysing the phagosomal membrane so as to avoid being consumed and degraded by lysosomal enzymes. They enter the cytosol in order to acquire the necessary nutrients, energy, and growth factors. These bacteria have a long generation time of about 8-10 hours in a human host. They grow relatively slowly in order to survive for extended periods of time within their host cell. One study showed a possibility of actin-based mobility that allowed the pathogen to spread to neighboring cells. (Walker et al., 2003)
Rickettsia rickettsii live and multiply in small- to medium-sized blood vessels in the human body, most often in the brain, skin, and the heart. They are able to grow in the cytoplasm or the nucleus of the host cell. When the bacteria replicate, this results in severe damage and often death of the cells in which it inhabits. During multiplication, blood leaks out into nearby tissues through holes in vessel walls. This process is known as hyperplasia where the endothelial cells and localized thrombus formation obstruct the flow of blood. Inflammatory cells also accumulate among the infected segments of blood vessels. (CDC, 2005)
Current research supports the response involving the epithelial cells damaging these blood vessels causing increased vascular permeability. Often times the leaking of the excess fluid from the blood vessels can impair organ function. These cells also produced reactive oxygen species that damage the infected cells through the lipid peroxidation of the host cell membrane. This creates an additional oxygen stress-mediated injury which eventually depletes some components of the host cell while levels of catalase increase to defend the cell against the dangerous reactive oxygen species. (Walker et al., 2003) This mechanism of injury to blood vessels was suggested by the morphological analogy of the lesion to immune complex vasculitis, the clinical similarity of Rocky Mountain Spotted Fever and serum sickness, the presence of circulating rickettsiae and antibodies, and the demonstration of immunoglobulin and the third component of complement in late lesions in experimentally infected guinea pigs. (Walker et al., 1977) This part of the cycle involving injury to blood vessels causes the rash traditionally associated with Rocky Mountain Spotted Fever, in addition to other symptoms including stupor and terminal shock. Death is often caused through excessive damage to the endothelial cells, resulting in the leaking of plasma, decrease in blood volume, and shock. (CDC, 2005)